

"Negative vaccination" by specific CD4 T cell tolerisation enhances virus-specific protective antibody responses
- PMID: 18000535
- PMCID: PMC2048666
- DOI: 10.1371/journal.pone.0001162
"Negative vaccination" by specific CD4 T cell tolerisation enhances virus-specific protective antibody responses
Abstract
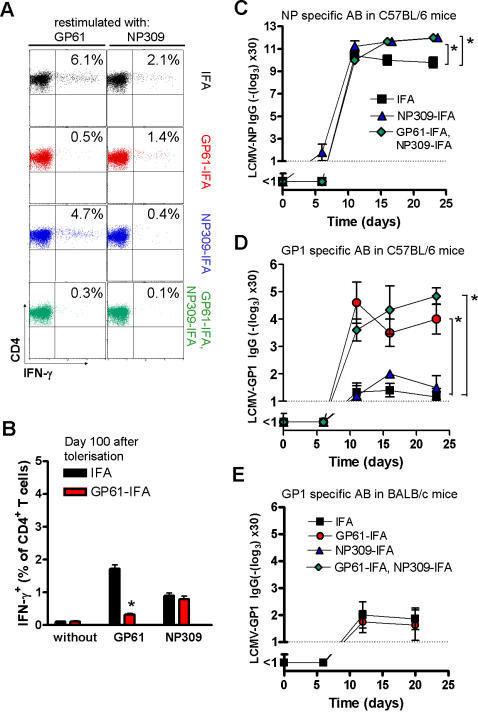
Background: Cooperation of CD4+ T helper cells with specific B cells is crucial for protective vaccination against pathogens by inducing long-lived neutralizing antibody responses. During infection with persistence-prone viruses, prolonged virus replication correlates with low neutralizing antibody responses. We recently described that a viral mutant of lymphocytic choriomeningitis virus (LCMV), which lacks a T helper epitope, counterintuitively induced an enhanced protective antibody response. Likewise, partial depletion of the CD4+ T cell compartment by using anti-CD4 antibodies enhanced protective antibodies.
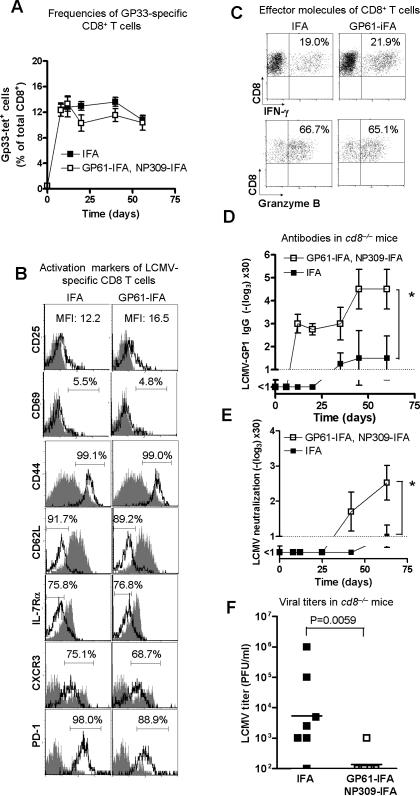
Principal findings: Here we have developed a protocol to selectively reduce the CD4+ T cell response against viral CD4+ T cell epitopes. We demonstrate that in vivo treatment with LCMV-derived MHC-II peptides induced non-responsiveness of specific CD4+ T cells without affecting CD4+ T cell reactivity towards other antigens. This was associated with accelerated virus-specific neutralizing IgG-antibody responses. In contrast to a complete absence of CD4+ T cell help, tolerisation did not impair CD8+ T cell responses.
Conclusions: This result reveals a novel "negative vaccination" strategy where specific CD4+ T cell unresponsiveness may be used to enhance the delayed protective antibody responses in chronic virus infections.
Conflict of interest statement
Figures


Similar articles
-
Virus-specific MHC-class II-restricted TCR-transgenic mice: effects on humoral and cellular immune responses after viral infection.Eur J Immunol. 1998 Jan;28(1):390-400. doi: 10.1002/(SICI)1521-4141(199801)28:01<390::AID-IMMU390>3.0.CO;2-O. Eur J Immunol. 1998. PMID: 9485218
-
CD4(+) T cells induced by a DNA vaccine: immunological consequences of epitope-specific lysosomal targeting.J Virol. 2001 Nov;75(21):10421-30. doi: 10.1128/JVI.75.21.10421-10430.2001. J Virol. 2001. PMID: 11581410 Free PMC article.
-
Impairment and delay of neutralizing antiviral antibody responses by virus-specific cytotoxic T cells.J Immunol. 1993 Nov 15;151(10):5408-15. J Immunol. 1993. PMID: 7693811
-
Analyzing Mouse B Cell Responses Specific to LCMV Infection.Methods Mol Biol. 2018;1707:15-38. doi: 10.1007/978-1-4939-7474-0_2. Methods Mol Biol. 2018. PMID: 29388097 Review.
-
T and B cell tolerance and responses to viral antigens in transgenic mice: implications for the pathogenesis of autoimmune versus immunopathological disease.Immunol Rev. 1991 Aug;122:133-71. doi: 10.1111/j.1600-065x.1991.tb00601.x. Immunol Rev. 1991. PMID: 1937540 Review.
Cited by
-
Influence of disulfide-stabilized structure on the specificity of helper T-cell and antibody responses to HIV envelope glycoprotein gp120.J Virol. 2010 Apr;84(7):3303-11. doi: 10.1128/JVI.02242-09. Epub 2010 Jan 20. J Virol. 2010. PMID: 20089653 Free PMC article.
-
The neutralization breadth of HIV-1 develops incrementally over four years and is associated with CD4+ T cell decline and high viral load during acute infection.J Virol. 2011 May;85(10):4828-40. doi: 10.1128/JVI.00198-11. Epub 2011 Mar 9. J Virol. 2011. PMID: 21389135 Free PMC article.
-
Cross-reactive broadly neutralizing antibodies: timing is everything.Front Immunol. 2012 Jul 20;3:215. doi: 10.3389/fimmu.2012.00215. eCollection 2012. Front Immunol. 2012. PMID: 22833745 Free PMC article.
-
Antibodies to the CD4-binding site of HIV-1 gp120 suppress gp120-specific CD4 T cell response while enhancing antibody response.Infect Agent Cancer. 2008 Jul 18;3:11. doi: 10.1186/1750-9378-3-11. Infect Agent Cancer. 2008. PMID: 18638381 Free PMC article.
-
MyD88 intrinsically regulates CD4 T-cell responses.J Virol. 2009 Feb;83(4):1625-34. doi: 10.1128/JVI.01770-08. Epub 2008 Dec 3. J Virol. 2009. PMID: 19052080 Free PMC article.
References
-
- Ada G. Vaccines and Vaccination. N Engl J Med. 2001;345:1042–53. - PubMed
-
- Koff WC, Johnson PR, Watkins DI, Burton DR, Lifson JD, et al. Hiv Vaccine Design: Insights from Live Attenuated Siv Vaccines. Nat Immunol. 2006;7:19–23. - PubMed
-
- Inchauspe G, Feinstone S. Development of a Hepatitis C Virus Vaccine. Clin Liver Dis. 2003;7:243–59, xi. - PubMed
-
- Mullen GE, Giersing BK, Ajose-Popoola O, Davis HL, Kothe C, et al. Enhancement of Functional Antibody Responses to Ama1-C1/Alhydrogel((R)), a Plasmodium Falciparum Malaria Vaccine, with Cpg Oligodeoxynucleotide. Vaccine. 2006;24:2497–505. - PubMed
-
- von Reyn CF. Tuberculosis Vaccine Trials. Lancet Infect Dis. 2005;5:132. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
