

Modeling the bicoid gradient: diffusion and reversible nuclear trapping of a stable protein
- PMID: 18001703
- PMCID: PMC2789574
- DOI: 10.1016/j.ydbio.2007.09.058
Modeling the bicoid gradient: diffusion and reversible nuclear trapping of a stable protein
Erratum in
- Dev Biol. 2008 Apr 15;316(2):548
Abstract
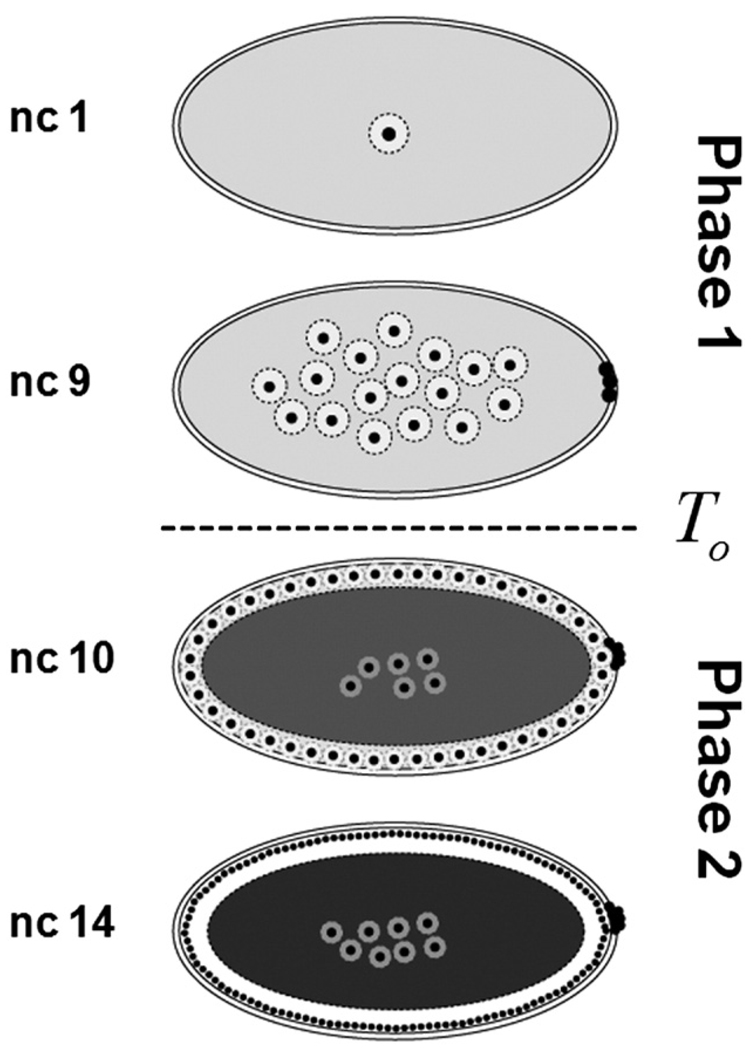
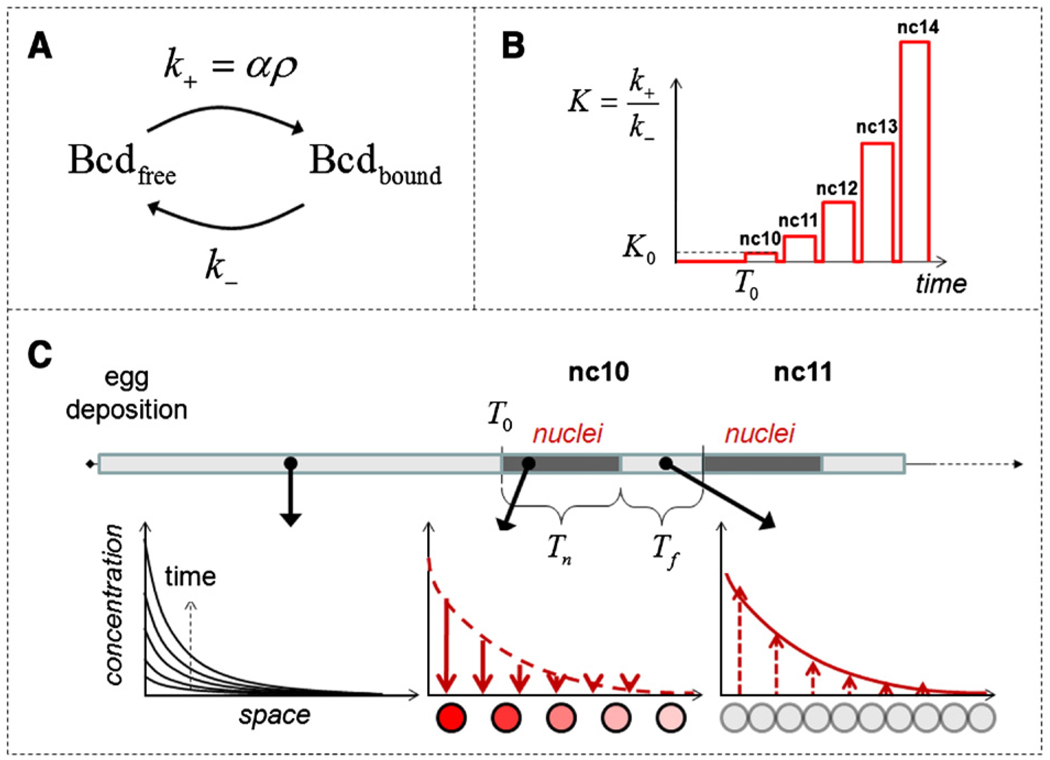
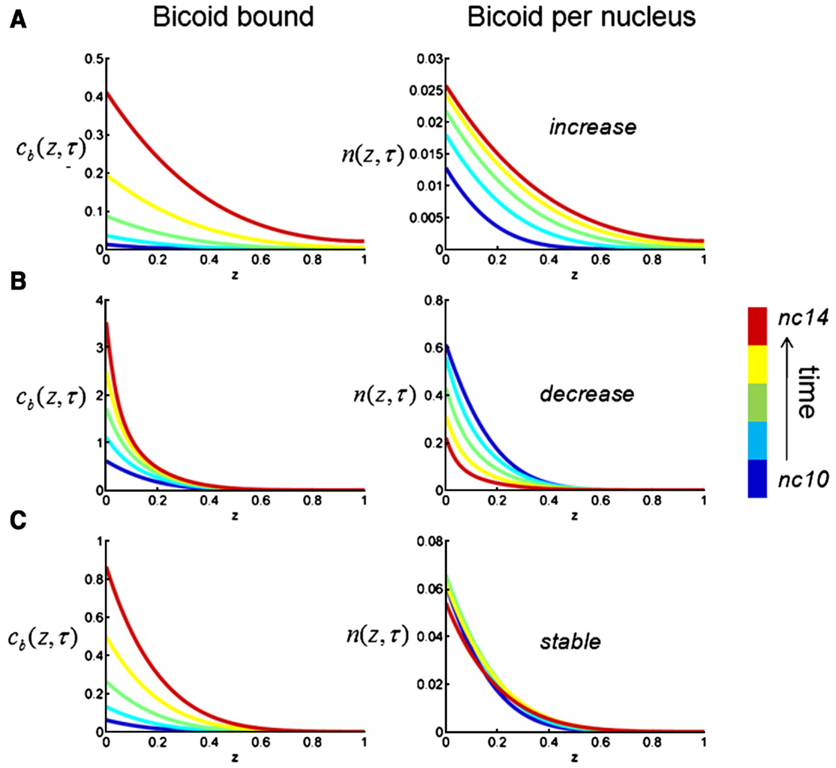
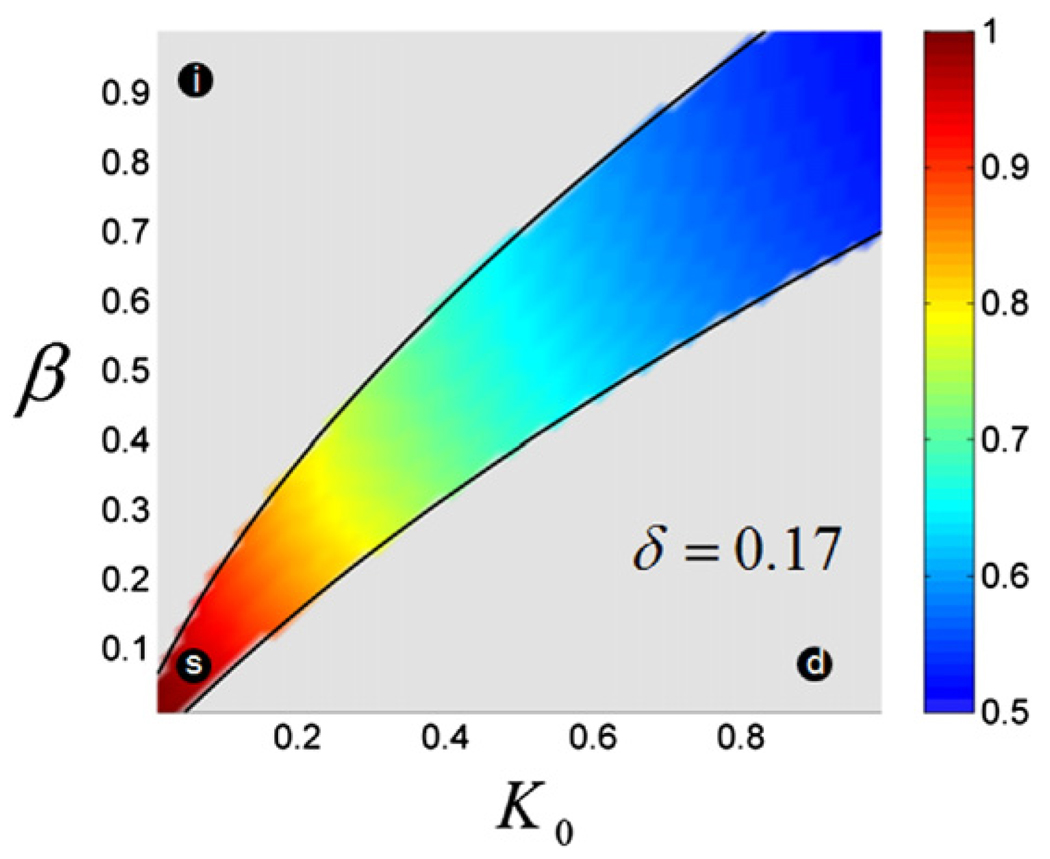
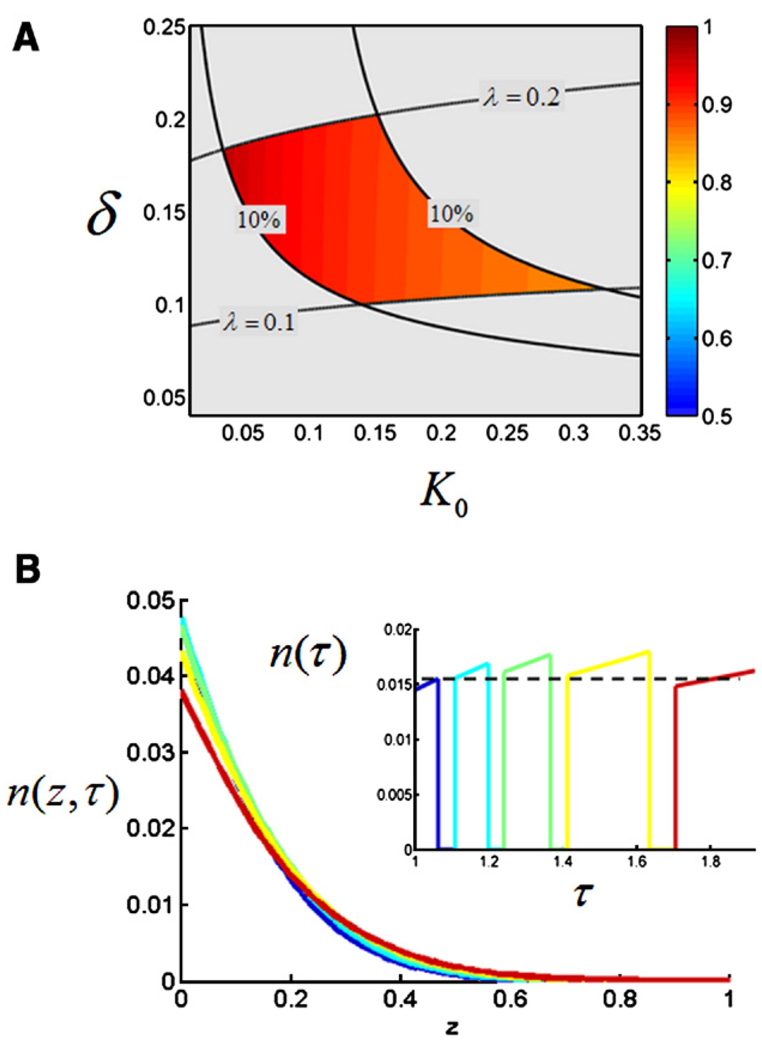
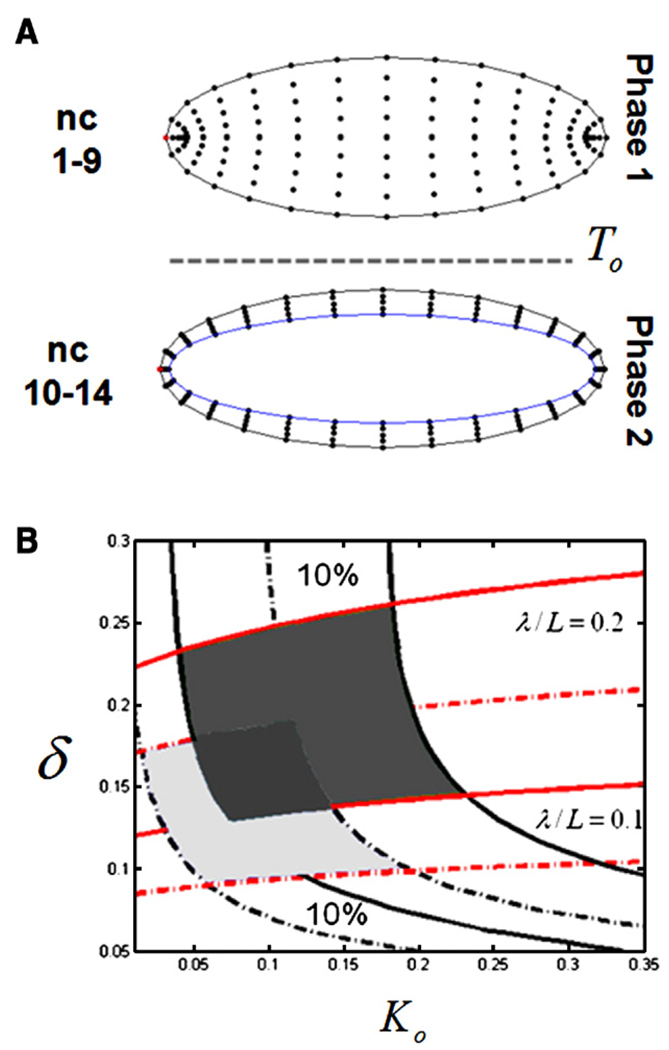
The Bicoid gradient in the Drosophila embryo provided the first example of a morphogen gradient studied at the molecular level. The exponential shape of the Bicoid gradient had always been interpreted within the framework of the localized production, diffusion, and degradation model. We propose an alternative mechanism, which assumes no Bicoid degradation. The medium where the Bicoid gradient is formed and interpreted is very dynamic. Most notably, the number of nuclei changes over three orders of magnitude from fertilization, when Bicoid synthesis is initiated, to nuclear cycle 14 when most of the measurements were taken. We demonstrate that a model based on Bicoid diffusion and nucleocytoplasmic shuttling in the presence of the growing number of nuclei can account for most of the properties of the Bicoid concentration profile. Consistent with experimental observations, the Bicoid gradient in our model is established before nuclei migrate to the periphery of the embryo and remains stable during subsequent nuclear divisions.
Figures
Similar articles
-
A precise Bicoid gradient is nonessential during cycles 11-13 for precise patterning in the Drosophila blastoderm.PLoS One. 2008;3(11):e3651. doi: 10.1371/journal.pone.0003651. Epub 2008 Nov 7. PLoS One. 2008. PMID: 18989373 Free PMC article.
-
The formation of the Bicoid morphogen gradient requires protein movement from anteriorly localized mRNA.PLoS Biol. 2011 Mar;9(3):e1000596. doi: 10.1371/journal.pbio.1000596. Epub 2011 Mar 1. PLoS Biol. 2011. PMID: 21390295 Free PMC article.
-
Determining the scale of the Bicoid morphogen gradient.Proc Natl Acad Sci U S A. 2009 Feb 10;106(6):1710-5. doi: 10.1073/pnas.0807655106. Epub 2009 Feb 3. Proc Natl Acad Sci U S A. 2009. PMID: 19190186 Free PMC article.
-
Seeing is believing: the bicoid morphogen gradient matures.Cell. 2004 Jan 23;116(2):143-52. doi: 10.1016/s0092-8674(04)00037-6. Cell. 2004. PMID: 14744427 Review.
-
A matter of time: Formation and interpretation of the Bicoid morphogen gradient.Curr Top Dev Biol. 2020;137:79-117. doi: 10.1016/bs.ctdb.2019.11.016. Epub 2019 Dec 27. Curr Top Dev Biol. 2020. PMID: 32143754 Review.
Cited by
-
Context-dependent transcriptional interpretation of mitogen activated protein kinase signaling in the Drosophila embryo.Chaos. 2013 Jun;23(2):025105. doi: 10.1063/1.4808157. Chaos. 2013. PMID: 23822503 Free PMC article.
-
Scaling of the Bicoid morphogen gradient by a volume-dependent production rate.Development. 2011 Jul;138(13):2741-9. doi: 10.1242/dev.064402. Epub 2011 May 25. Development. 2011. PMID: 21613328 Free PMC article.
-
Semi-permeable Diffusion Barriers Enhance Patterning Robustness in the C. elegans Germline.Dev Cell. 2015 Nov 23;35(4):405-17. doi: 10.1016/j.devcel.2015.10.027. Dev Cell. 2015. PMID: 26609956 Free PMC article.
-
A compartmental model for the bicoid gradient.Dev Biol. 2010 Sep 1;345(1):12-7. doi: 10.1016/j.ydbio.2010.05.491. Epub 2010 May 24. Dev Biol. 2010. PMID: 20580703 Free PMC article.
-
Modelling the Bicoid gradient.Development. 2010 Jul;137(14):2253-64. doi: 10.1242/dev.032409. Development. 2010. PMID: 20570935 Free PMC article. Review.
References
-
- Abramowitz M, Stegun IA. Handbook of Mathematical Tables with Formulas, Graphs, and Mathematical Tables. Washington: Dover Pubns; 1964.
-
- Bressloff PC, Earnshaw BA. Diffusion-trapping model of receptor trafficking in dendrites. Phys. Rev. 2007:E 75. - PubMed
-
- Casanova J, Struhl G. The torso receptor localizes as well as transduces the spatial signal specifying terminal body pattern in Drosophila. Nature. 1993;362:152–155. - PubMed
-
- Chen Y, Struhl G. Dual roles for patched in sequestering and transducing hedgehog. Cell. 1996;87:553–563. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
