

Disease effects on reproduction can cause population cycles in seasonal environments
- PMID: 18005128
- PMCID: PMC2408661
- DOI: 10.1111/j.1365-2656.2007.01328.x
Disease effects on reproduction can cause population cycles in seasonal environments
Abstract
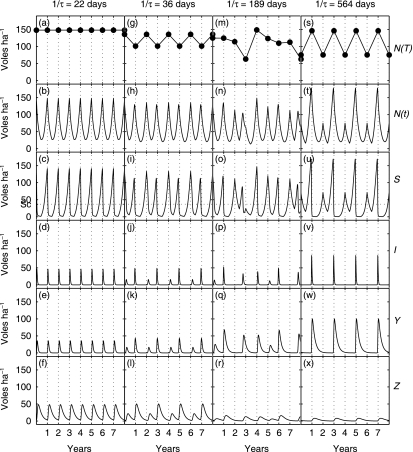
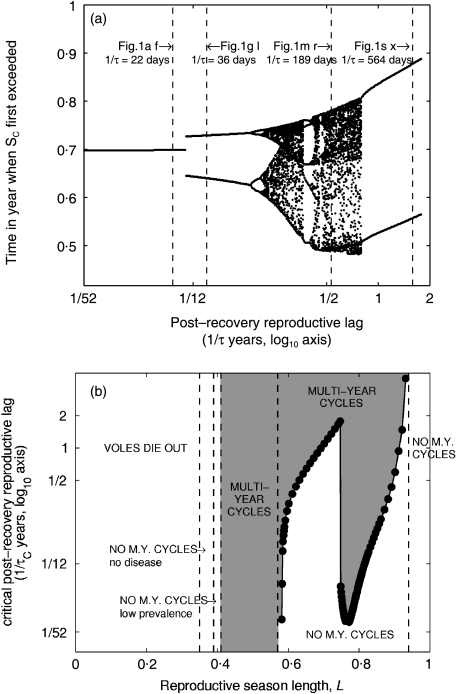
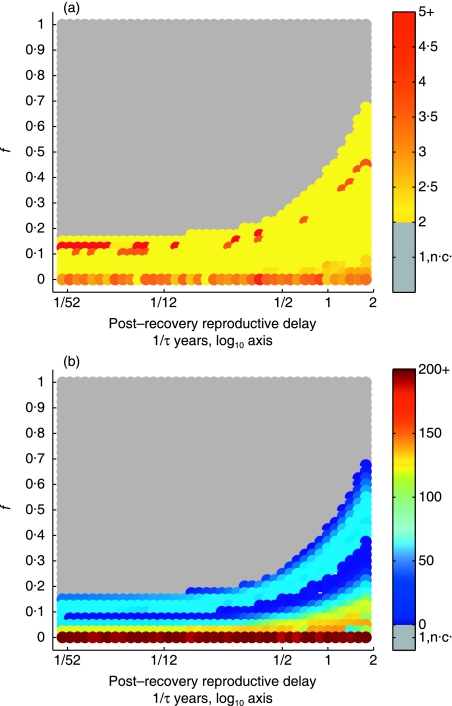
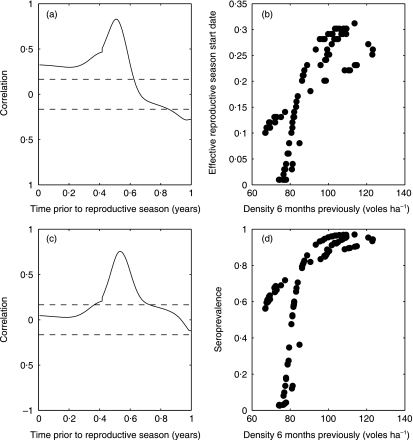
1. Recent studies of rodent populations have demonstrated that certain parasites can cause juveniles to delay maturation until the next reproductive season. Furthermore, a variety of parasites may share the same host, and evidence is beginning to accumulate showing nonindependent effects of different infections. 2. We investigated the consequences for host population dynamics of a disease-induced period of no reproduction, and a chronic reduction in fecundity following recovery from infection (such as may be induced by secondary infections) using a modified SIR (susceptible, infected, recovered) model. We also included a seasonally varying birth rate as recent studies have demonstrated that seasonally varying parameters can have important effects on long-term host-parasite dynamics. We investigated the model predictions using parameters derived from five different cyclic rodent populations. 3. Delayed and reduced fecundity following recovery from infection have no effect on the ability of the disease to regulate the host population in the model as they have no effect on the basic reproductive rate. However, these factors can influence the long-term dynamics including whether or not they exhibit multiyear cycles. 4. The model predicts disease-induced multiyear cycles for a wide range of realistic parameter values. Host populations that recover relatively slowly following a disease-induced population crash are more likely to show multiyear cycles. Diseases for which the period of infection is brief, but full recovery of reproductive function is relatively slow, could generate large amplitude multiyear cycles of several years in length. Chronically reduced fecundity following recovery can also induce multiyear cycles, in support of previous theoretical studies. 5. When parameterized for cowpox virus in the cyclic field vole populations (Microtus agrestis) of Kielder Forest (northern England), the model predicts that the disease must chronically reduce host fecundity by more than 70%, following recovery from infection, for it to induce multiyear cycles. When the model predicts quasi-periodic multiyear cycles it also predicts that seroprevalence and the effective date of onset of the reproductive season are delayed density-dependent, two phenomena that have been recorded in the field.
Figures

References
-
- Altizer S, Dobson A, Hosseini P, Hudson P, Pascual M, Rohani P. Seasonality and the dynamics of infectious diseases. Ecology Letters. 2006;9:467–484. - PubMed
-
- Anderson RM, May RM. The population-dynamics of micro-parasites and their invertebrate hosts. Philosophical Transactions of the Royal Society of London, Series B, Biological Sciences. 1981;291:451–524.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
