

Heparan sulfate proteoglycans provide a signal to Plasmodium sporozoites to stop migrating and productively invade host cells
- PMID: 18005753
- PMCID: PMC2117360
- DOI: 10.1016/j.chom.2007.10.002
Heparan sulfate proteoglycans provide a signal to Plasmodium sporozoites to stop migrating and productively invade host cells
Abstract
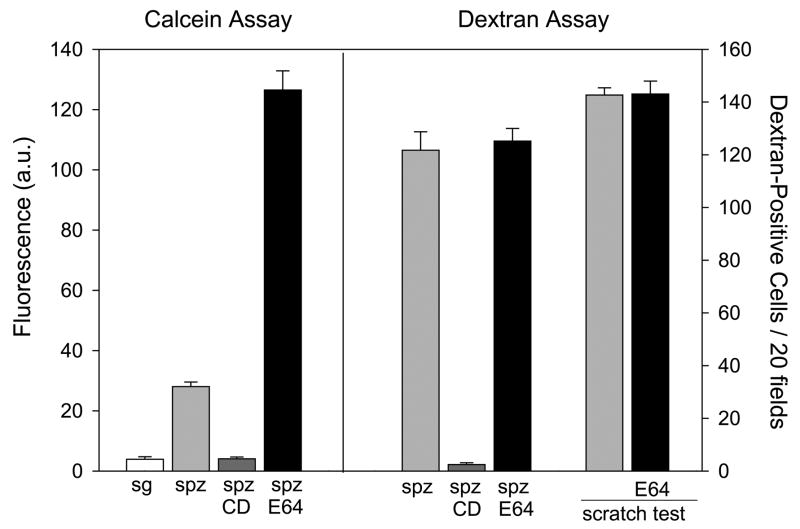
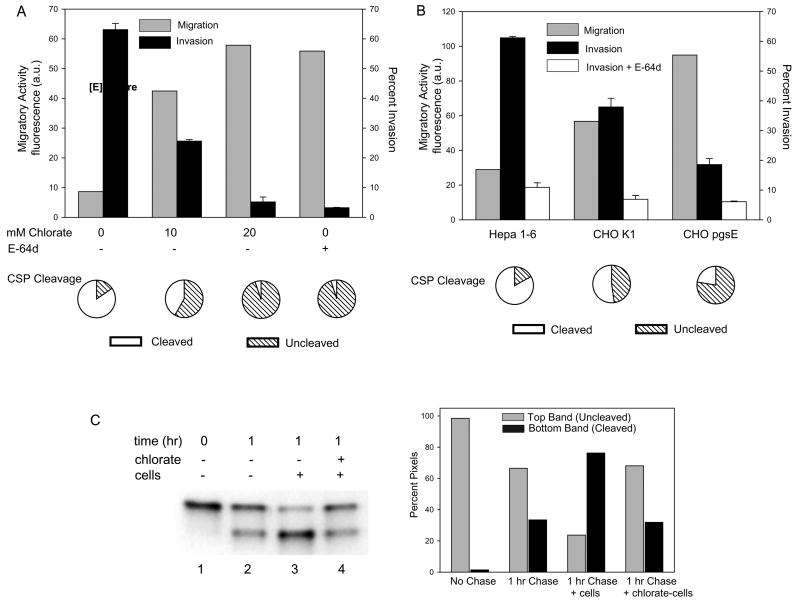
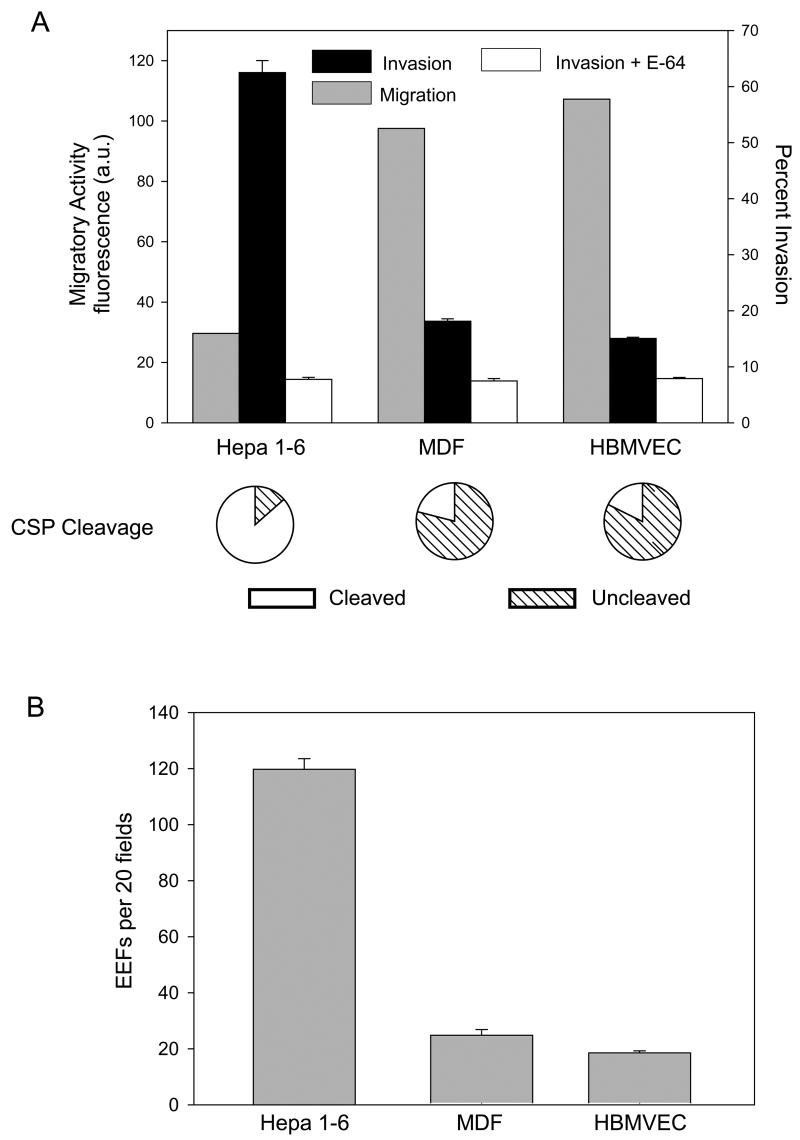
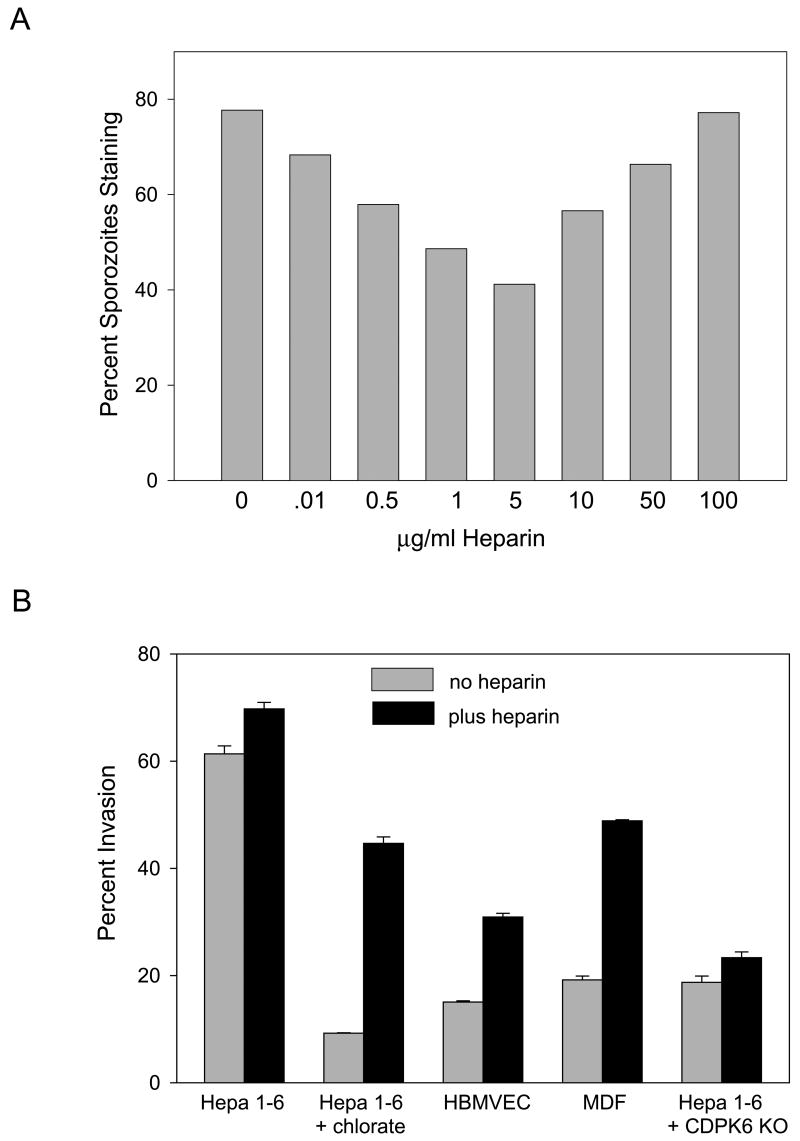
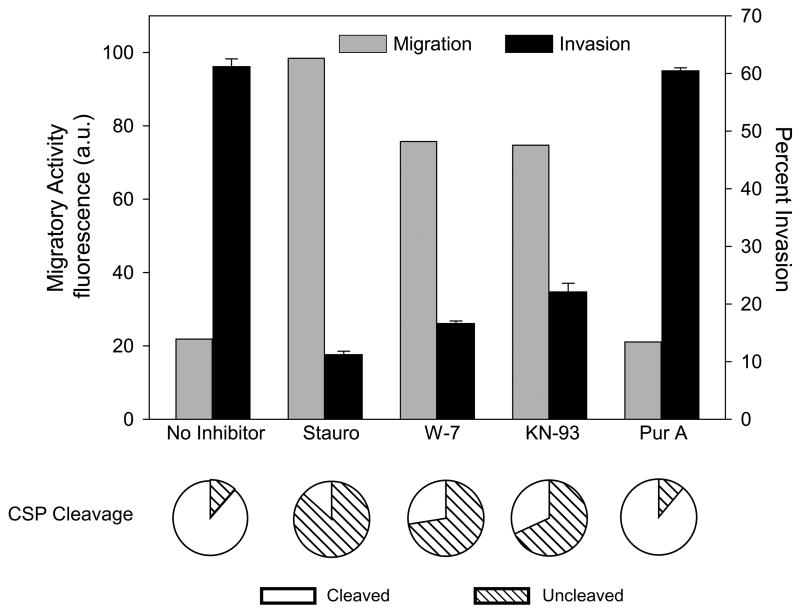
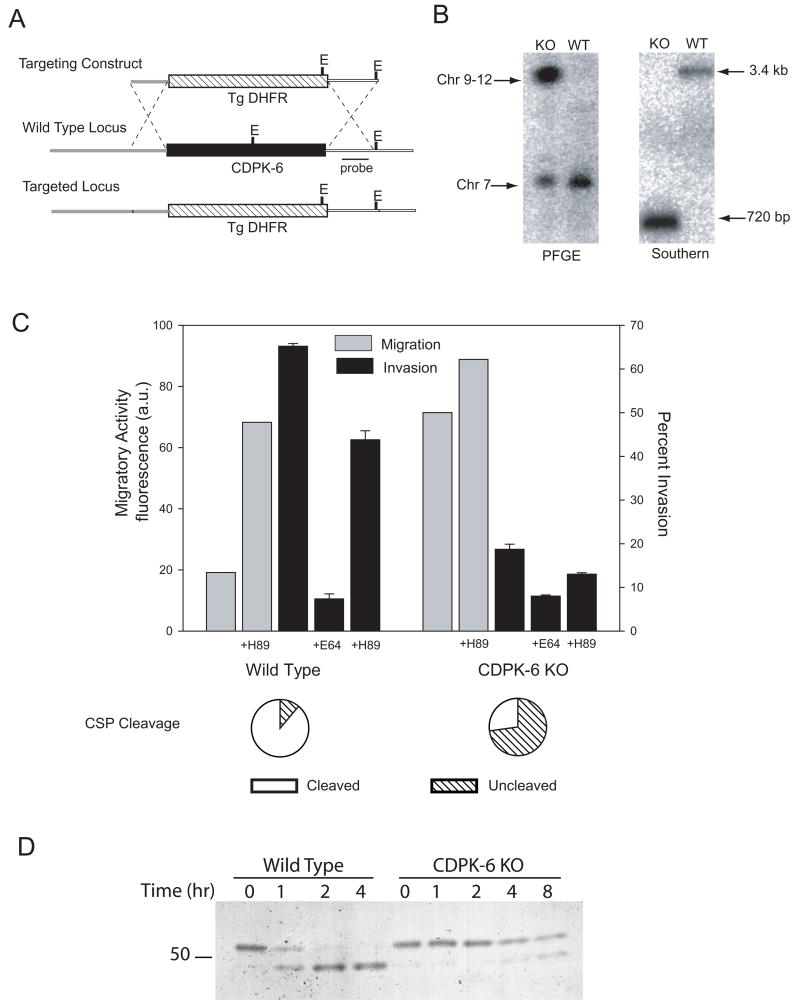
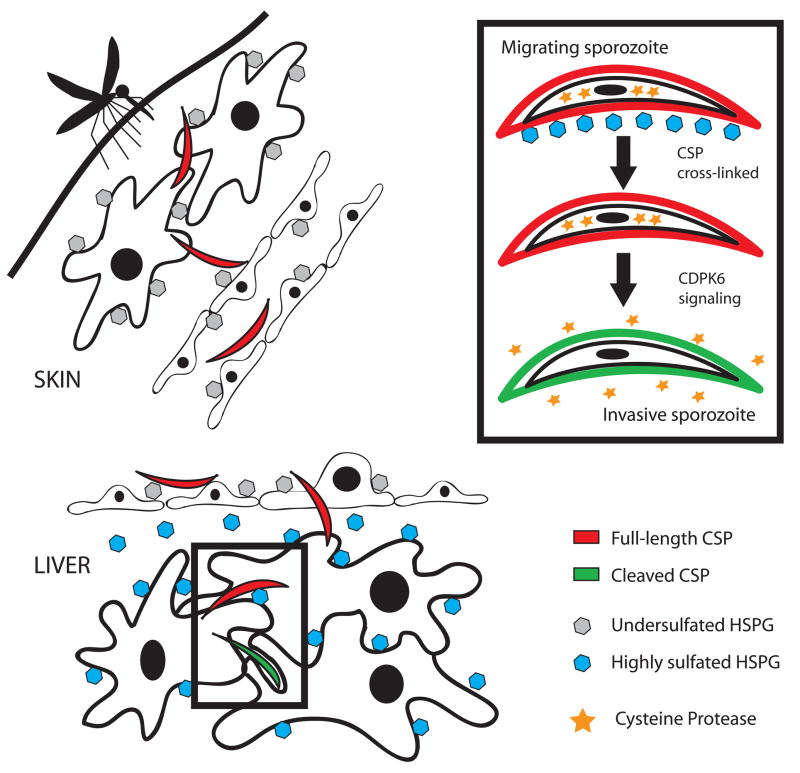
Malaria infection is initiated when Anopheles mosquitoes inject Plasmodium sporozoites into the skin. Sporozoites subsequently reach the liver, invading and developing within hepatocytes. Sporozoites contact and traverse many cell types as they migrate from skin to liver; however, the mechanism by which they switch from a migratory mode to an invasive mode is unclear. Here, we show that sporozoites of the rodent malaria parasite Plasmodium berghei use the sulfation level of host heparan sulfate proteoglycans (HSPGs) to navigate within the mammalian host. Sporozoites migrate through cells expressing low-sulfated HSPGs, such as those in skin and endothelium, while highly sulfated HSPGs of hepatocytes activate sporozoites for invasion. A calcium-dependent protein kinase is critical for the switch to an invasive phenotype, a process accompanied by proteolytic cleavage of the sporozoite's major surface protein. These findings explain how sporozoites retain their infectivity for an organ that is far from their site of entry.
Figures
Comment in
-
To migrate or to invade: those are the options.Cell Host Microbe. 2007 Nov 15;2(5):286-8. doi: 10.1016/j.chom.2007.10.008. Cell Host Microbe. 2007. PMID: 18005748 Review.
References
-
- Amino R, Thiberge S, Martin B, Celli S, Shorte S, Frischknecht F, Menard R. Quantitative imaging of Plasmodium transmission from mosquito to mammal. Nat Med. 2006;12:220–224. - PubMed
-
- Bame KJ, Esko JD. Undersulfated heparan sulfate in a Chinese Hamster Ovary cell mutant defective in heparan sulfate N-sulfotransferase. J Biol Chem. 1989;264:8059–8065. - PubMed
-
- Bhanot P, Schauer K, Coppens I, Nussenzweig V. A surface phospholipase is involved in the migration of Plasmodium sporozoites through cells. J Biol Chem. 2005;280:6752–6760. - PubMed
-
- Billker O, Dechamps S, Tewari R, Wenig G, Franke-Fayard B, Brinkmann V. Calcium and a calcium-dependent protein kinase regulate gamete formation and mosquito transmission in a malaria parasite. Cell. 2004;117:503–514. - PubMed
-
- Carruthers VB, Sibley LD. Mobilization of intracellular calcium stimulates microneme discharge in Toxoplasma gondii. Mol Microbiol. 1999;31:421–428. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
