

Adiponectin activates adenosine monophosphate-activated protein kinase and decreases luteinizing hormone secretion in LbetaT2 gonadotropes
- PMID: 18006641
- PMCID: PMC2262174
- DOI: 10.1210/me.2007-0330
Adiponectin activates adenosine monophosphate-activated protein kinase and decreases luteinizing hormone secretion in LbetaT2 gonadotropes
Abstract
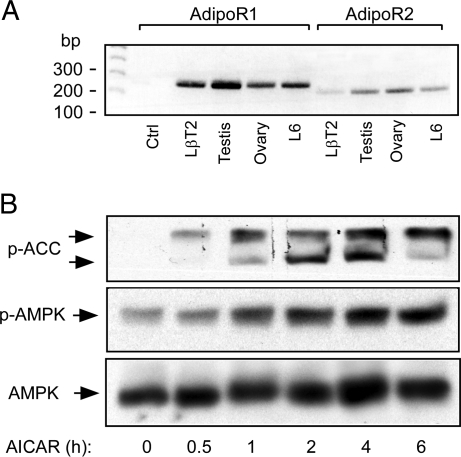
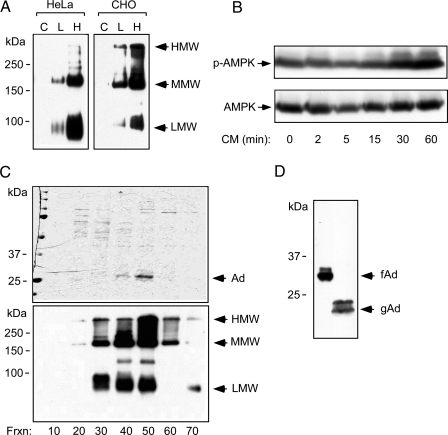
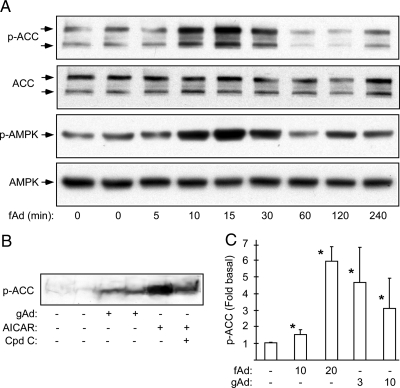
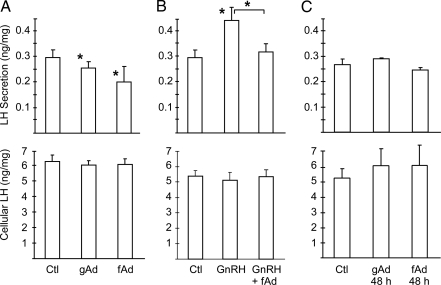
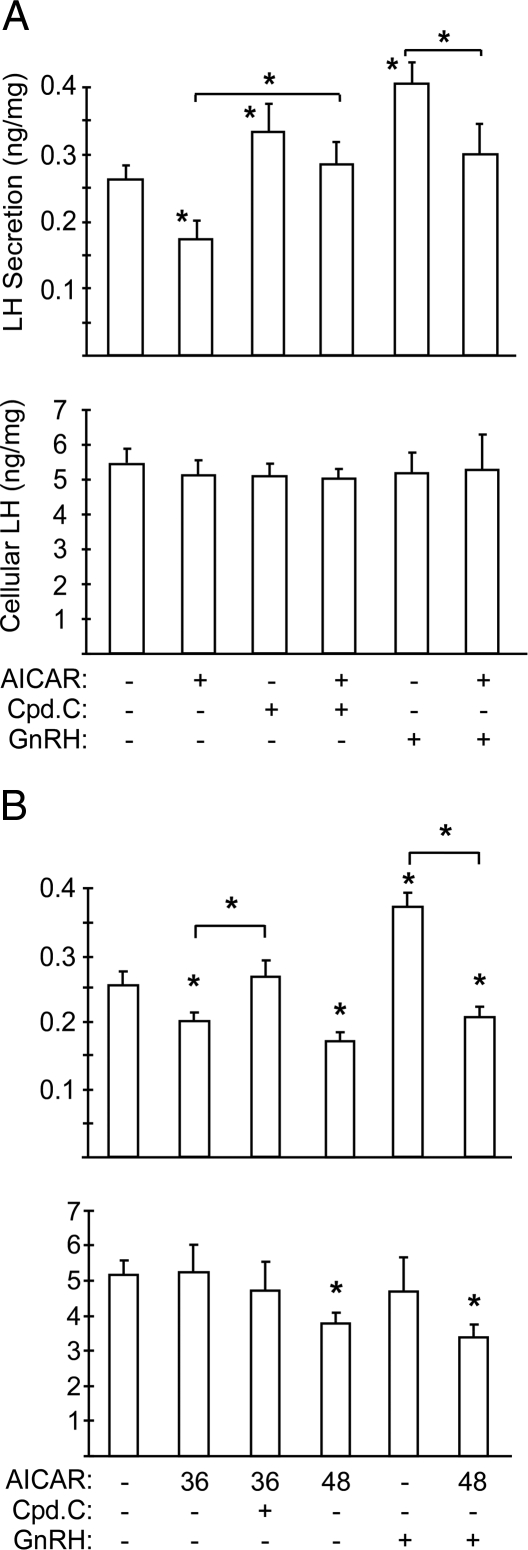
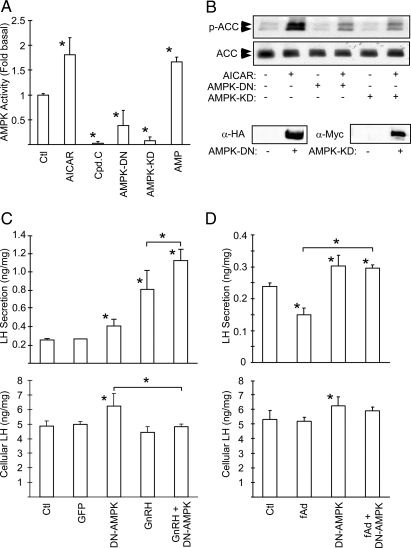
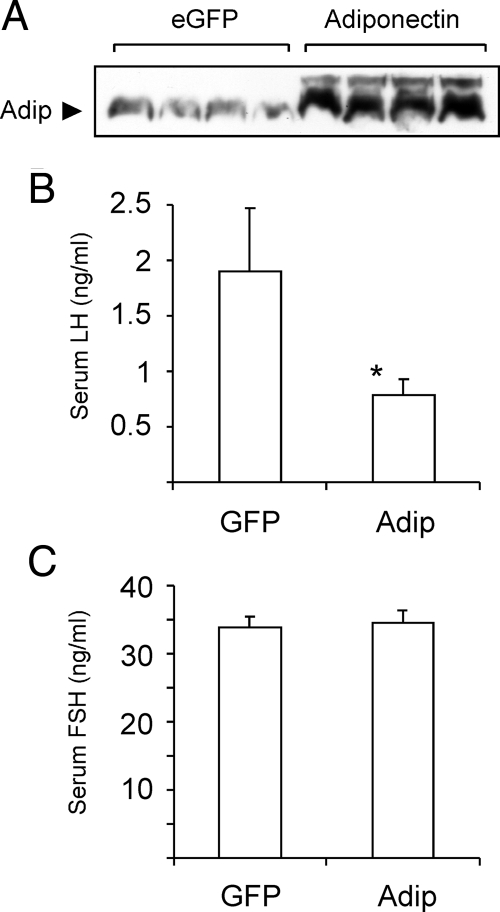
Metabolic dysregulation is associated with reproductive disorders, but the underlying mechanisms are not clearly understood. Adiponectin is an adipocyte-derived secretory factor that improves insulin sensitivity. Results from animal models indicate that overexpression of adiponectin impairs female fertility. We hypothesized that adiponectin regulates reproduction by altering the hypothalamic-pituitary axis. Mouse LbetaT2 immortalized gonadotrope cells express both adiponectin receptors 1 and 2. Adiponectin increases phosphorylation of AMP-activated protein kinase (AMPK), a downstream target of adiponectin receptors, and reduces basal and GnRH-stimulated LH secretion, acutely. The repression of LH secretion can be mimicked by 5-aminoimidazole-4-carboxamide-1-beta-riboside, an AMP analog, suggesting the involvement of AMPK. A dominant-negative AMPK mutant or compound C, a selective AMPK inhibitor, potentiates basal LH secretion and abolishes the inhibitory effect of adiponectin. Chronic activation of AMPK by 5-aminoimidazole-4-carboxamide-1-beta-riboside decreases cellular LH levels, and expression of dominant-negative AMPK increases cellular LH levels, suggesting a second effect of AMPK to regulate LH synthesis. Lastly, intravenous injection of an adenovirus expressing adiponectin into male mice reduces serum LH levels without changing FSH levels. In conclusion, our results suggest that adiponectin decreases LH secretion in pituitary gonadotropes in an AMPK-dependent manner.
Figures
References
-
- Schneider JE 2004 Energy balance and reproduction. Physiol Behav 81:289–317 - PubMed
-
- Wade GN, Jones JE 2004 Neuroendocrinology of nutritional infertility. Am J Physiol 287:R1277–R1296 - PubMed
-
- Mitchell M, Armstrong DT, Robker RL, Norman RJ 2005 Adipokines: implications for female fertility and obesity. Reproduction (Cambridge, England) 130:583–597 - PubMed
-
- Trujillo ME, Scherer PE 2006 Adipose tissue-derived factors: impact on health and disease. Endocr Rev 27:762–778 - PubMed
-
- Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM 1994 Positional cloning of the mouse obese gene and its human homologue. Nature 372:425–432 - PubMed
