

Molecular basis for agonist selectivity and activation of the orphan bombesin receptor subtype 3 receptor
- PMID: 18006692
- PMCID: PMC2593135
- DOI: 10.1124/jpet.107.132332
Molecular basis for agonist selectivity and activation of the orphan bombesin receptor subtype 3 receptor
Abstract
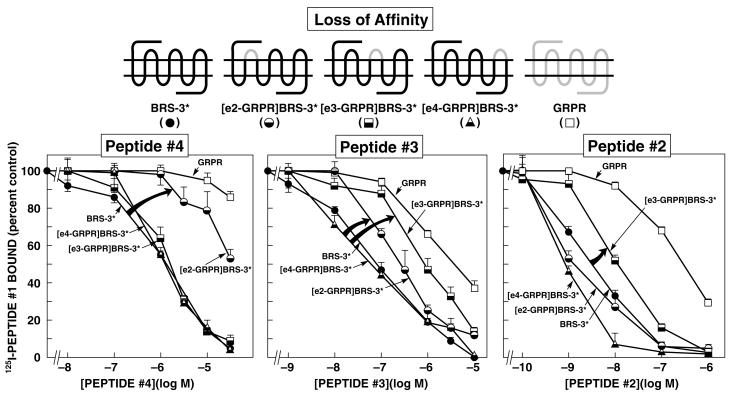
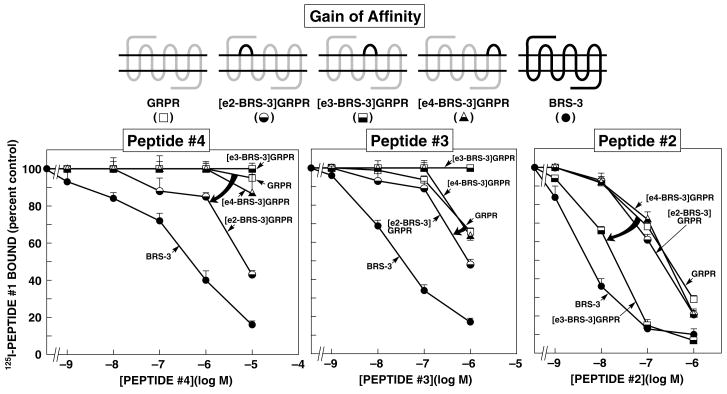
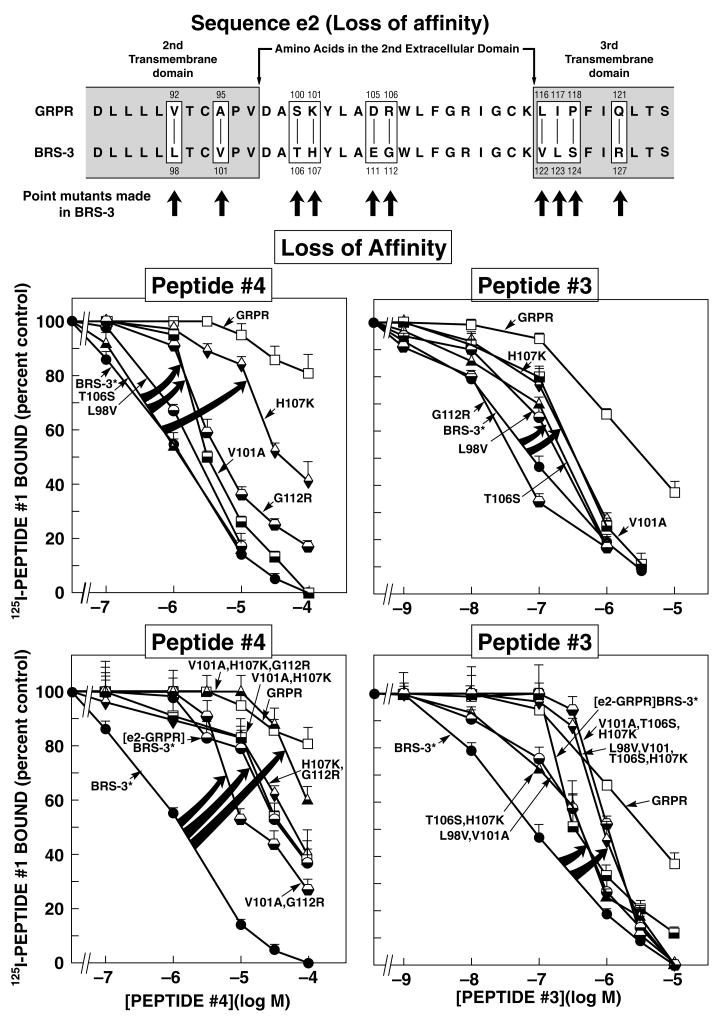
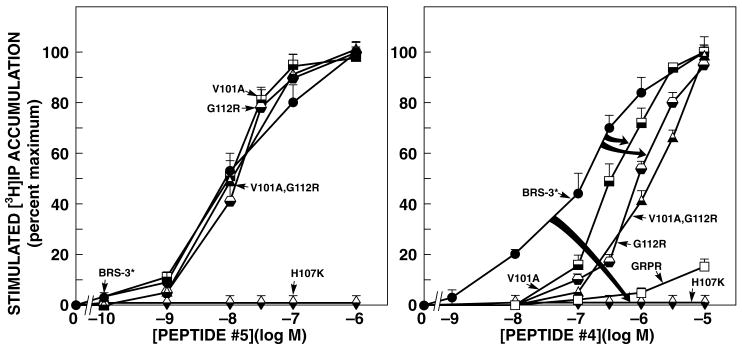
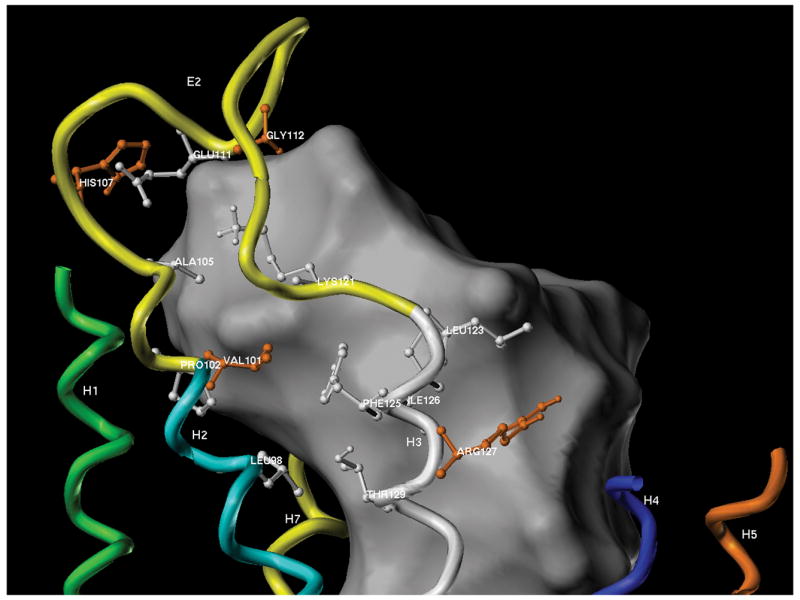
Bombesin receptor subtype (BRS)-3, a G-protein-coupled orphan receptor, shares 51% identity with the mammalian bombesin (Bn) receptor for gastrin-releasing peptide. There is increasing interest in BRS-3 because it is important in energy metabolism, glucose control, motility, and tumor growth. BRS-3 has low affinity for all Bn-related peptides; however, recently synthetic high-affinity agonists, [d-Tyr(6)/d-Phe(6),betaAla(11),Phe(13),Nle(14)]Bn-(6-14), were described, but they are nonselective for BRS-3 over other Bn receptors. Based on these peptides, three BRS-3-selective ligands were developed: peptide 2, [d-Tyr(6)(R)-3-amino-propionic acid(11),Phe(13),Nle(14)]Bn(6-14); peptide 3, [d-Tyr(6),(R)-Apa(11),4Cl-Phe(13),Nle(14)]Bn(6-14); and peptide 4, acetyl-Phe-Trp-Ala-His-(tBzl)-piperidine-3 carboxylic acid-Gly-Arg-NH(2). Their molecular determinants of selectivity/high affinity for BRS-3 are unknown. To address this, we used a chimeric/site mutagenesis approach. Substitution of extracellular domain 2 (EC2) of BRS-3 by the comparable gastrin-releasing peptide receptor (GRPR) domain decreased 26-, 4-, and 0-fold affinity for peptides 4, 3, and 2. Substitution of EC3 decreased affinity 4-, 11-, and 0-fold affinity for peptides 2 to 4. Ten-point mutations in the EC2 and adjacent transmembrane regions (TM2) 2 and 3 of BRS-3 were made. His107 (EC2-BRS-3) for lysine (H107K) (EC2-GRPR) decreased affinity (25- and 0-fold) for peptides 4 and 1; however, it could not be activated by either peptide. Its combination with Val101 (TM2), Gly112 (EC2), and Arg127 (TM3) resulted in complete loss-of-affinity of peptide 4. Receptor-modeling showed that each of these residues face inward and are within 4 A of the binding pocket. These results demonstrate that Val101, His107, Gly112, and Arg127 in the EC2/adjacent upper TMs of BRS-3 are critical for the high BRS3 selectivity of peptide 4. His107 in EC2 is essential for BRS-3 activation, suggesting amino-aromatic ligand/receptor interactions with peptide 4 are critical for both binding and activation. Furthermore, these result demonstrate that even though these three BRS-3-selective agonists were developed from the same template peptide, [d-Phe(6),betaAla(11),Phe(13),Nle(14)]Bn-(6-14), their molecular determinants of selectivity/high affinity varied considerably.
Figures
References
-
- Akeson M, Sainz E, Mantey SA, Jensen RT, Battey JF. Identification of four amino acids in the gastrin-releasing peptide C receptor that are required for high affinity agonist binding. J Biol Chem. 1997;272:17405–17409. - PubMed
-
- Benya RV, Akeson M, Mrozinski J, Jensen RT, Battey JF. Internalization of the gastrin-releasing peptide receptor is mediated by phospholipase C-dependent and -independent processes. Mol Pharmacol. 1994;46:495–501. - PubMed
-
- Boyle RG, Humphries J, Mitchell T, Showell GA, Apaya R, Iijima H, Shimada H, Arai T, Ueno H, Usui Y, Sakaki T, Wada E, Wada K. The design of a new potent and selective ligand for the orphan bombesin receptor subtype 3 (BRS3) J Pept Sci. 2005;11:136–141. - PubMed
-
- Couvineau A, Rouyer-Fessard C, Maoret JJ, Gaudin P, Nicole P, Laburthe M. Vasoactive intestinal peptide (VIP)1 receptor. Three nonadjacent amino acids are responsible for species selectivity with respect to recognition of peptide histidine isoleucinemide. J Biol Chem. 1996;271:12795–12800. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
