

SM-protein-controlled ER-associated degradation discriminates between different SNAREs
- PMID: 18007658
- PMCID: PMC2267232
- DOI: 10.1038/sj.embor.7401105
SM-protein-controlled ER-associated degradation discriminates between different SNAREs
Abstract
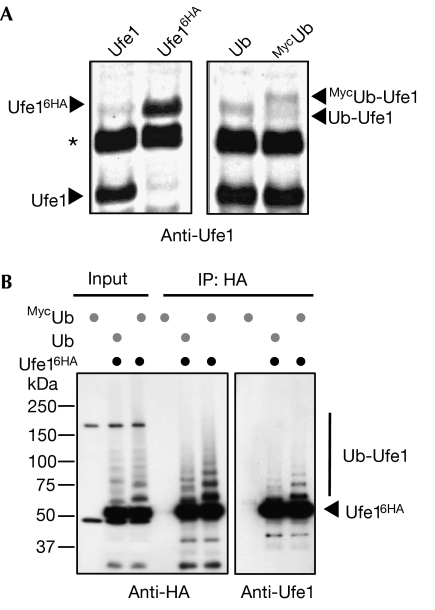
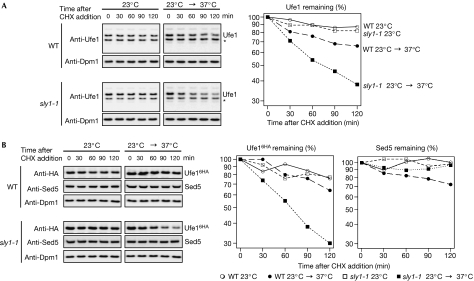
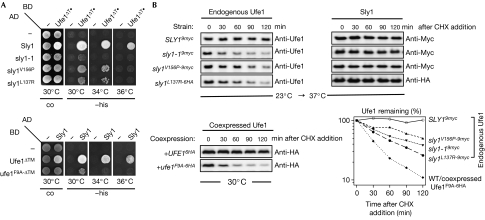
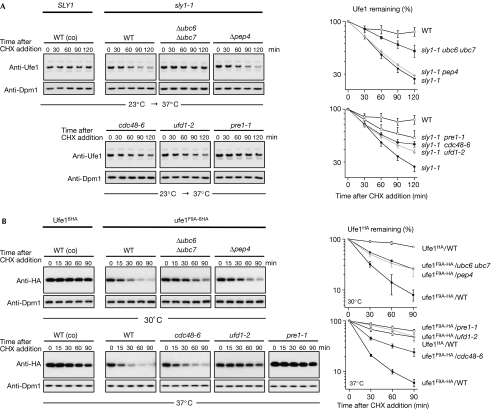
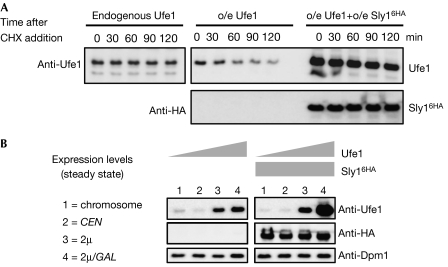
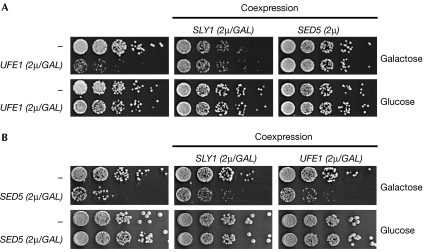
Endoplasmic reticulum (ER)-associated degradation (ERAD) is a specialized activity of the ubiquitin-proteasome system that is involved in clearing the ER of aberrant proteins and regulating the levels of specific ER-resident proteins. Here we show that the yeast ER-SNARE Ufe1, a syntaxin (Qa-SNARE) involved in ER membrane fusion and retrograde transport from the Golgi to the ER, is prone to degradation by an ERAD-like mechanism. Notably, Ufe1 is protected against degradation through binding to Sly1, a known SNARE regulator of the Sec1-Munc18 (SM) protein family. This mechanism is specific for Ufe1, as the stability of another Sly1 partner, the Golgi Qa-SNARE Sed5, is not influenced by Sly1 interaction. Thus, our findings identify Sly1 as a discriminating regulator of SNARE levels and indicate that Sly1-controlled ERAD might regulate the balance between different Qa-SNARE proteins.
Figures
References
-
- Biederer T, Volkwein C, Sommer T (1997) Role of Cue1p in ubiquitination and degradation at the ER surface. Science 278: 1806–1809 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
