

Review
doi: 10.1038/nri2199.
Protective immune mechanisms in helminth infection
Affiliations
- PMID: 18007680
- PMCID: PMC2258092
- DOI: 10.1038/nri2199
Item in Clipboard
Review
Protective immune mechanisms in helminth infection
Nat Rev Immunol.
2007 Dec.
Abstract
Important insights have recently been gained in our understanding of how host immune responses mediate resistance to parasitic helminths and control associated pathological responses. Although similar cells and cytokines are evoked in response to infection by helminths as diverse as nematodes and schistosomes, the components of the response that mediate protection are dependent on the particular parasite. In this Review, we examine recent findings regarding the mechanisms of protection in helminth infections that have been elucidated in murine models and discuss the implications of these findings in terms of future therapies.
Figures
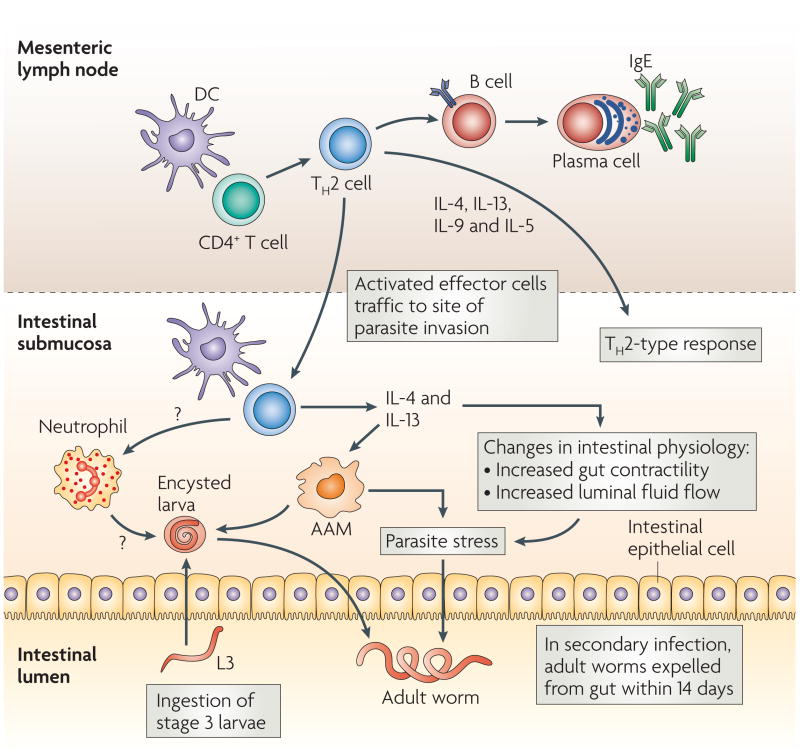
Infective third-stage larvae (L3) are ingested by the host. They then travel to the duodenum, invade the epithelia, and reside in the submucosa for 8 days, after which they re-enter the lumen of the duodenum as adult nematodes. Primary infections become established and chronic, but can be cleared by antihelminthic drug treatment. Challenge (secondary) infections are naturally cleared by the host by day 14 post-infection, making this an excellent model of protective memory T helper 2 (TH2)-type responses. Parasite antigens are presented to CD4+ T cells in mesenteric lymph nodes and other gut-associated lymphoid tissues, driving the induction of TH2 effector cells. These cells exert their effector functions through the production of a number of cytokines, including interleukin-4 (IL-4), IL-13, IL-9 and IL-5. TH2 cells induce B-cell immunoglobulin class-switching to IgE. Shortly after activation, TH2 effector cells migrate to the site of parasite residence in the submucosa. Within several days, a distinct immune-cell infiltrate appears which can damage the larval parasite after secondary, but not primary, inoculation. The infiltrate following secondary inoculation includes TH2 cells, dendritic cells (DCs), neutrophils and alternatively activated macrophages (AAMs). The TH2 cytokines IL-4 and IL-13 might also facilitate expulsion of adult parasites in the lumen by inducing changes in intestinal physiology.
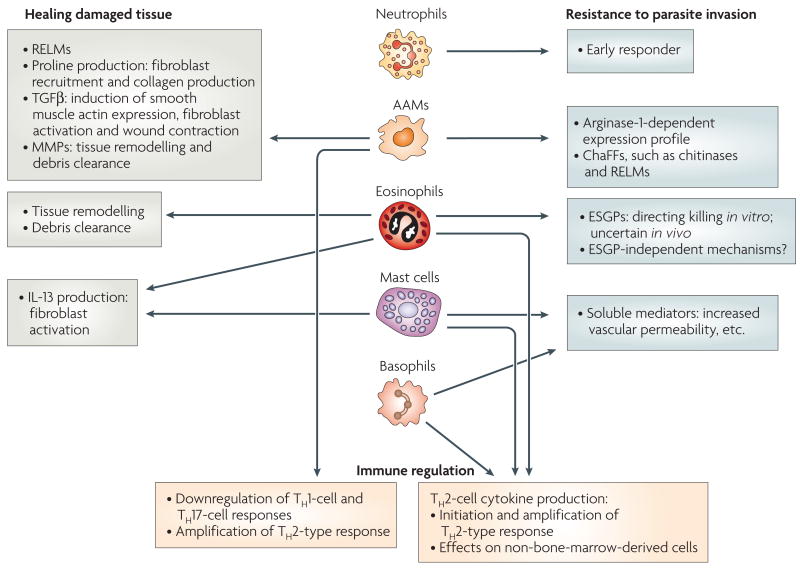
In general, the effector functions of innate immune cells during protective T helper 2 (TH2)-type responses can include healing damaged tissue, mediating resistance to parasite invasion and regulating the immune response. Alternatively activated macrophages (AAMs) and eosinophils might contribute to the healing of tissue damage caused by invasive helminth parasites through the production of factors that initiate and promote the remodelling and restructuring of tissue. AAMs, eosinophils, basophils, mast cells and neutrophils can contribute to resistance to parasite invasion. AAMs and eosinophils can directly stress tissue-dwelling parasites. Basophils and mast cells produce soluble mediators (including leukotrienes, prostaglandins and histamine) that promote luminal fluid flow, nerve stimulation and gut contractility. Neutrophils are early responders to invasive helminths, and are localized adjacent to the parasites. AAMs can regulate the immune response by downregulating TH1 and TH17 cells and promoting TH2 cells. Eosinophils, basophils and mast cells can produce TH2-type cytokines including interleukin-4 (IL-4) and IL-13, and thereby amplify the TH2-type response. ChaFFs, chitinase and FIZZ family members; ESGPs, eosinophil secondary granule proteins; MMPs, matrix metalloproteases; RELMs, resistin-like molecules, TGFβ, transforming growth factor-β.
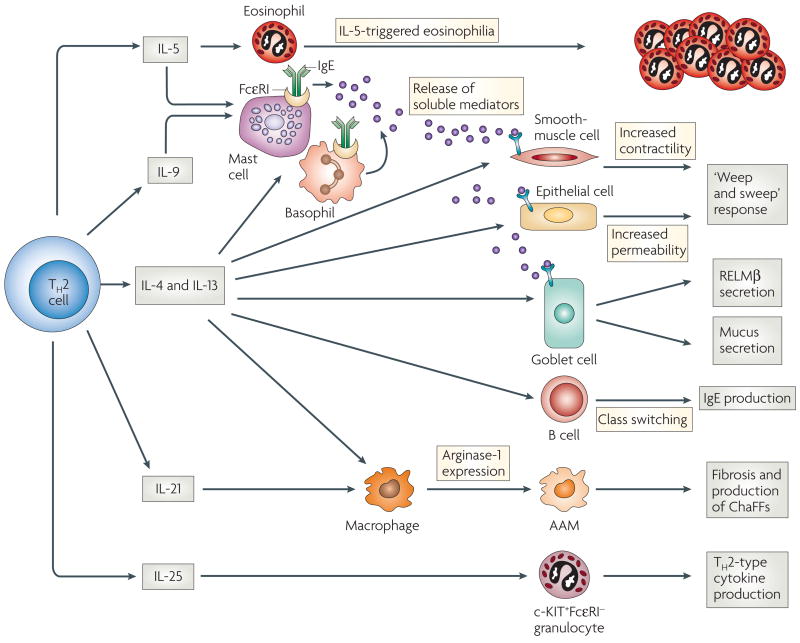
T helper 2 (TH2) cells orchestrate the immune response primarily through the production of cytokines in the lymph nodes and periphery. Interleukin-5 (IL-5) triggers eosinophilia, and in conjunction with IL-4, IL-9, and IL-13, and the crosslinking of FcεRIs (high-affinity Fc receptors for IgE), can result in enhanced mast-cell and basophil development and release of mediators. IL-4 and IL-13 stimulate increased smooth-muscle-cell contractility, increased intestinal permeability and elevated goblet-cell mucous secretion. IL-4 and IL-13 also enhance responsiveness of these cell types to mast-cell-derived mediators. Collectively, these effects can contribute to the ‘weep and sweep’ response to intestinal helminths. IL-4, in conjunction with other signals, can induce class switching in B cells, leading to IgE production. IL-4, IL-13 and IL-21, can drive development of alternatively activated macrophages (AAMs), leading to upregulation of arginase-1 expression, and in some cases this might lead to fibrosis, as in chronic schistosomiasis. IL-25 expression stimulates a c-KIT+FcεRI− population to migrate to lymph nodes and upregulate TH2-type cytokine mRNAs. It is unclear whether IL-25 is a TH2-type cytokine, or whether it is expressed by a distinct TH-cell lineage. IL-21, also produced by TH17 cells, is instrumental in the development of a TH17-like response (not shown). ChaFFs, chitinase and FIZZ family members; RELM, resistin-like molecule.
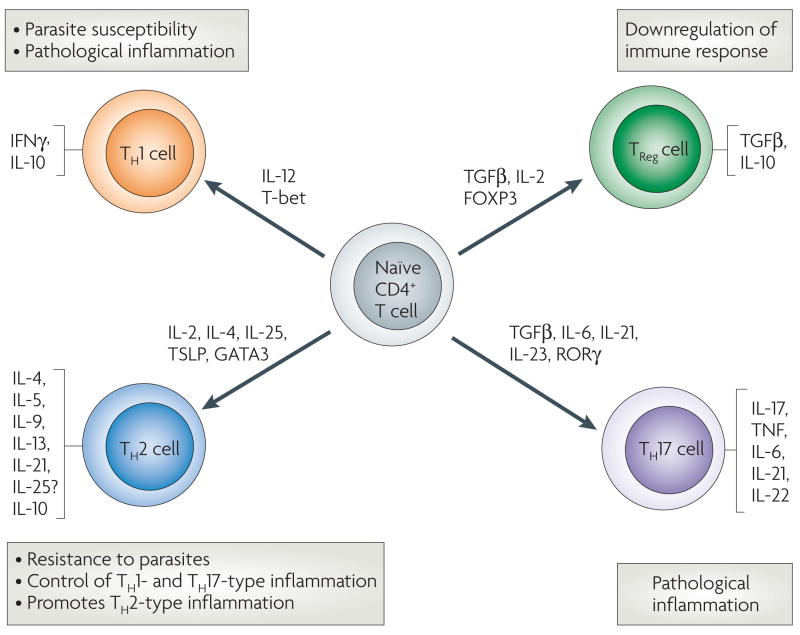
Naive CD4+ T cells can differentiate into several different types of effector and regulatory cells during helminth infection. Specific cytokines and transcription factors contribute to differentiation and expansion of these cell populations, and their differential activation plays a major role in determining whether an immune response will contribute to host protection or pathological inflammation. Exposure to interleukin-12 (IL-12) produced by antigen-presenting dendritic cells (DCs) induces T-bet expression, driving T helper 1 (TH1)-cell differentiation. These cells produce interferon-γ (IFNγ), and are unable to clear helminth parasites. The circumstances leading to TH2-cell differentiation are not as clear but recently, IL-2, IL-25 and thymic stromal lymphopoietin (TSLP), and associated transcription factors, have been implicated. TH2 cells produce a panel of cytokines that contribute to antihelminth immunity by instructing innate bone-marrow- and non-bone-marrow-derived cells, which in turn can instruct and amplify TH2 cells. Strong TH2-type responses often lead to increased resistance to parasites, as seen in Heligmosomoides polygyrus infection, or control of harmful inflammation mediated by TH1 cells and TH17 cells, as in schistosomiasis. However, chronic potent TH2-cell responses can also lead to harmful TH2-type inflammatory responses including fibrosis. Expression of the transcription factor forkhead box P3 (FOXP3) along with the cytokines transforming growth factor-β (TGFβ) and IL-2, leads to differentiation of regulatory T cells (TReg cells), which can inhibit the immune response. Several factors, including retinoic-acid-receptor-related orphan receptor-γ (RORγ), TGFβ, IL-6, IL-21 and IL-23, have been shown to promote TH17-cell differentiation, and these effector cells can mediate harmful inflammation. IL-10, which is no longer considered to be exclusively a TH2-type cytokine, can be produced by TH1, TH2, and TRegcells and downregulates both TH1- and TH2-type responses. TNF, tumour-necrosis factor.
References
-
- WHO. World Health Report. World Health Organization; Geneva, Switzerland: 1999.
-
- Capron A, Riveau G, Capron M, Trottein F. Schistosomes: the road from host-parasite interactions to vaccines in clinical trials. Trends Parasitol. 2005;21:143–149. - PubMed
-
- Goud GN, et al. Expression of the Necator americanus hookworm larval antigen Na-ASP-2 in Pichia pastoris and purification of the recombinant protein for use in human clinical trials. Vaccine. 2005;23:4754–4764. - PubMed
-
- Fallon PG, Mangan NE. Suppression of TH2-type allergic reactions by helminth infection. Nature Rev Immunol. 2007;7:220–230. - PubMed
-
- Wilson MS, Maizels RM. Regulation of allergy and autoimmunity in helminth infection. Clin Rev Allergy Immunol. 2004;26:35–50. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
