

Nanosize titanium dioxide stimulates reactive oxygen species in brain microglia and damages neurons in vitro
- PMID: 18007996
- PMCID: PMC2072833
- DOI: 10.1289/ehp.10216
Nanosize titanium dioxide stimulates reactive oxygen species in brain microglia and damages neurons in vitro
Abstract
Background: Titanium dioxide is a widely used nanomaterial whose photo-reactivity suggests that it could damage biological targets (e.g., brain) through oxidative stress (OS).
Objectives: Brain cultures of immortalized mouse microglia (BV2), rat dopaminergic (DA) neurons (N27), and primary cultures of embryonic rat striatum, were exposed to Degussa P25, a commercially available TiO(2) nanomaterial. Physical properties of P25 were measured under conditions that paralleled biological measures.
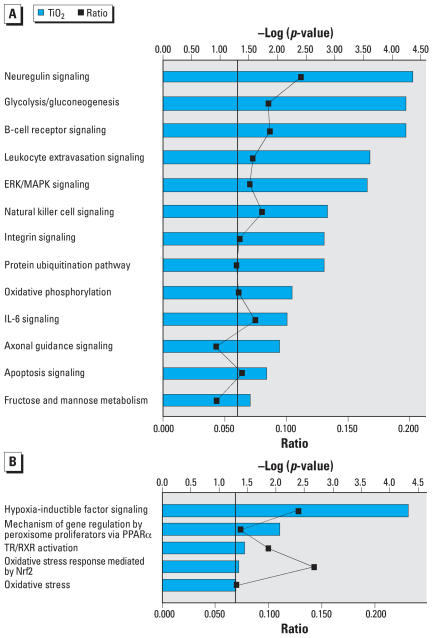
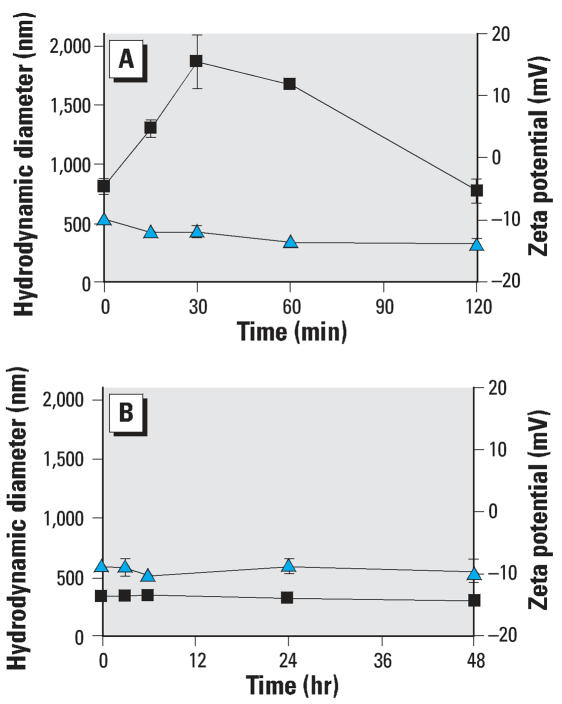
Findings: P25 rapidly aggregated in physiological buffer (800-1,900 nm; 25 degrees C) and exposure media (approximately 330 nm; 37 degrees C), and maintained a negative zeta potential in both buffer (-12.2 +/- 1.6 mV) and media (-9.1 +/- 1.2 mV). BV2 microglia exposed to P25 (2.5-120 ppm) responded with an immediate and prolonged release of reactive oxygen species (ROS). Hoechst nuclear stain was reduced after 24-hr (>or=100 ppm) and 48-hr (>or=2.5 ppm) exposure. Microarray analysis on P25-exposed BV2 microglia indicated up-regulation of inflammatory, apoptotic, and cell cycling pathways and down-regulation of energy metabolism. P25 (2.5-120 ppm) stimulated increases of intracellular ATP and caspase 3/7 activity in isolated N27 neurons (24-48 hr) but did not produce cytotoxicity after 72-hr exposure. Primary cultures of rat striatum exposed to P25 (5 ppm) showed a reduction of immunohistochemically stained neurons and microscopic evidence of neuronal apoptosis after 6-hr exposure. These findings indicate that P25 stimulates ROS in BV2 microglia and is nontoxic to isolated N27 neurons. However, P25 rapidly damages neurons at low concentrations in complex brain cultures, plausibly though microglial generated ROS.
Keywords: BV2; P25; environmental nanotoxicity; neurotoxicity; oxidative stress; titanium dioxide.
Figures

 resulting from interference with the mitochondria’s ETC was measured with MitoSOX Red. BV2 microglia, incubated in 2 μM (10 min, 37°C) showed a delayed but significant increase in fluorescence after 30-min exposure to ≥100 ppm P25. (C) Significant increases of caspase 3/7 activity were first seen by 6 hr in response to ≥40 ppm P25 and remained at this level for 24 hr. (D) Apoptotic loss of nuclear material, as measured with Hoechst stain, was first noted after 24 hr in response to P25 (≥100 ppm) and involved all concentrations by 48 hr.
resulting from interference with the mitochondria’s ETC was measured with MitoSOX Red. BV2 microglia, incubated in 2 μM (10 min, 37°C) showed a delayed but significant increase in fluorescence after 30-min exposure to ≥100 ppm P25. (C) Significant increases of caspase 3/7 activity were first seen by 6 hr in response to ≥40 ppm P25 and remained at this level for 24 hr. (D) Apoptotic loss of nuclear material, as measured with Hoechst stain, was first noted after 24 hr in response to P25 (≥100 ppm) and involved all concentrations by 48 hr.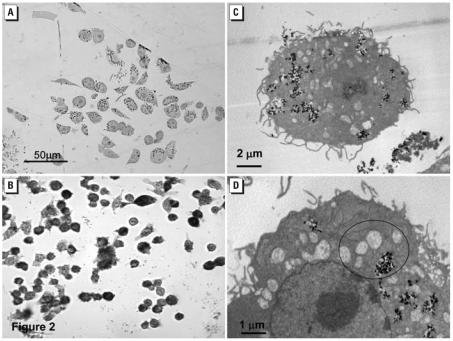





References
-
- Adams LK, Lyon DY, McIntosh A, Alvarez PJ. Comparative toxicity of nano-scale TiO2, SiO2 and ZnO water suspensions. Water Sci Technol. 2006;54:327–334. - PubMed
-
- Afaq F, Abidi P, Matin R, Rahman Q. Cytotoxicity, pro-oxidant effects and antioxidant depletion in rat lung alveolar macrophages exposed to ultrafine titanium dioxide. J Appl Toxicol. 1998;18:307–312. - PubMed
-
- Affymetrix Inc. Affymetrix Technical Manual. 2004. [[accessed 2 October 2007]]. Available: www.expressionanalysis.comm.
-
- Artuch R, Colome C, Vilaseca MA, Pineda M, Campistol J. Ubiquinone: metabolism and functions. Ubiquinone deficiency and its implication in mitochondrial encephalopathies. Treatment with ubiquinone. Rev Neurol. 1999;29:59–63. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials