

Experimental design for efficient identification of gene regulatory networks using sparse Bayesian models
- PMID: 18021391
- PMCID: PMC2233642
- DOI: 10.1186/1752-0509-1-51
Experimental design for efficient identification of gene regulatory networks using sparse Bayesian models
Abstract
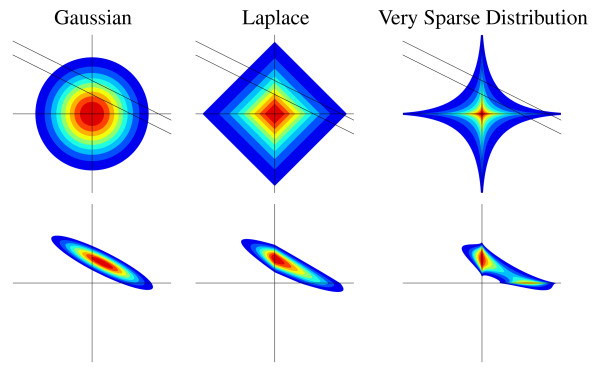
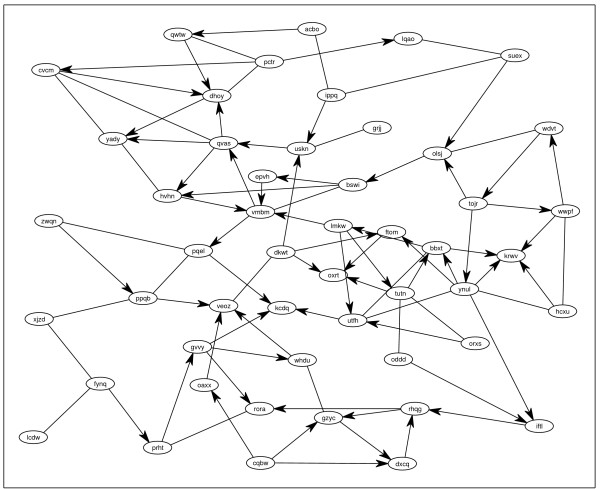
Background: Identifying large gene regulatory networks is an important task, while the acquisition of data through perturbation experiments (e.g., gene switches, RNAi, heterozygotes) is expensive. It is thus desirable to use an identification method that effectively incorporates available prior knowledge - such as sparse connectivity - and that allows to design experiments such that maximal information is gained from each one.
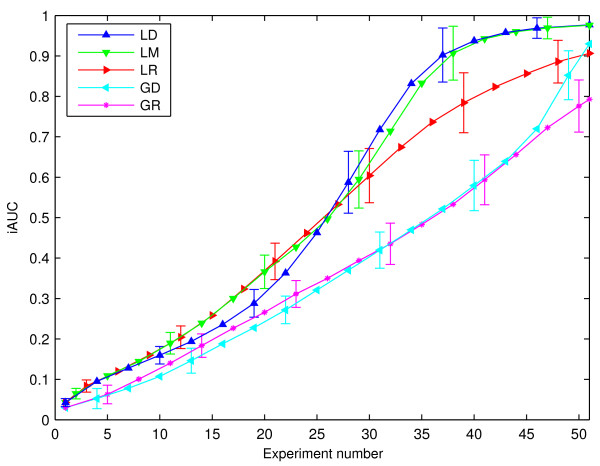
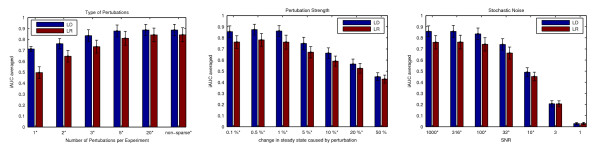
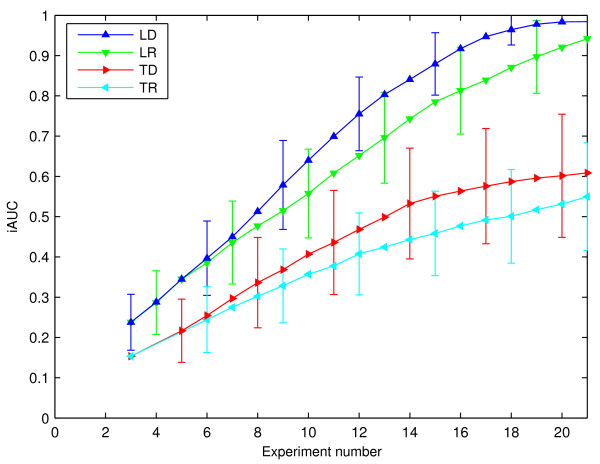
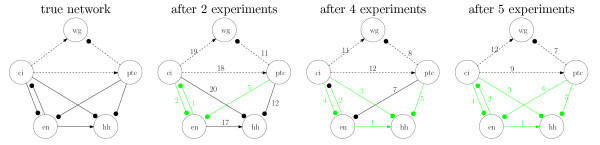
Results: Our main contributions are twofold: a method for consistent inference of network structure is provided, incorporating prior knowledge about sparse connectivity. The algorithm is time efficient and robust to violations of model assumptions. Moreover, we show how to use it for optimal experimental design, reducing the number of required experiments substantially. We employ sparse linear models, and show how to perform full Bayesian inference for these. We not only estimate a single maximum likelihood network, but compute a posterior distribution over networks, using a novel variant of the expectation propagation method. The representation of uncertainty enables us to do effective experimental design in a standard statistical setting: experiments are selected such that the experiments are maximally informative.
Conclusion: Few methods have addressed the design issue so far. Compared to the most well-known one, our method is more transparent, and is shown to perform qualitatively superior. In the former, hard and unrealistic constraints have to be placed on the network structure for mere computational tractability, while such are not required in our method. We demonstrate reconstruction and optimal experimental design capabilities on tasks generated from realistic non-linear network simulators. The methods described in the paper are available as a Matlab package athttp://www.kyb.tuebingen.mpg.de/sparselinearmodel.
Figures
References
MeSH terms
LinkOut - more resources
Full Text Sources
