

The [FeFe] hydrogenase of Nyctotherus ovalis has a chimeric origin
- PMID: 18021395
- PMCID: PMC2216082
- DOI: 10.1186/1471-2148-7-230
The [FeFe] hydrogenase of Nyctotherus ovalis has a chimeric origin
Abstract
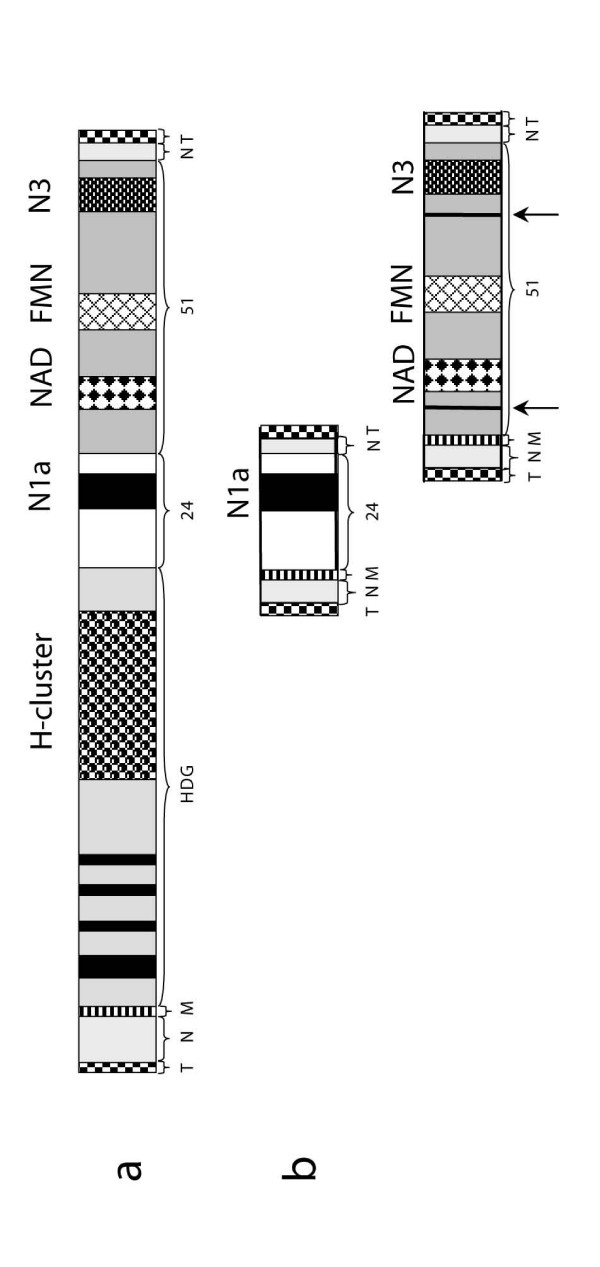
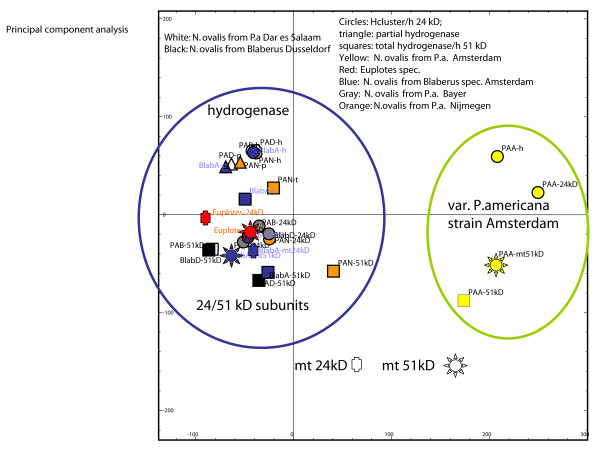
Background: The hydrogenosomes of the anaerobic ciliate Nyctotherus ovalis show how mitochondria can evolve into hydrogenosomes because they possess a mitochondrial genome and parts of an electron-transport chain on the one hand, and a hydrogenase on the other hand. The hydrogenase permits direct reoxidation of NADH because it consists of a [FeFe] hydrogenase module that is fused to two modules, which are homologous to the 24 kDa and the 51 kDa subunits of a mitochondrial complex I.
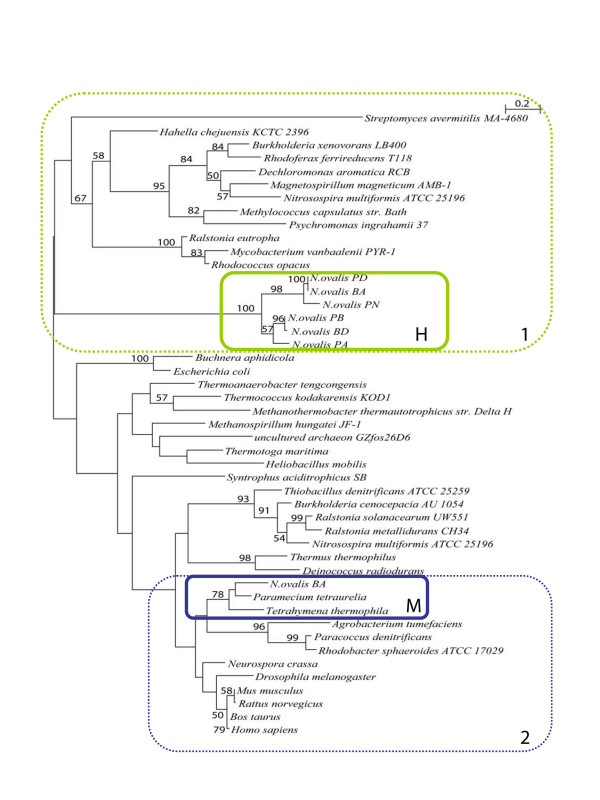
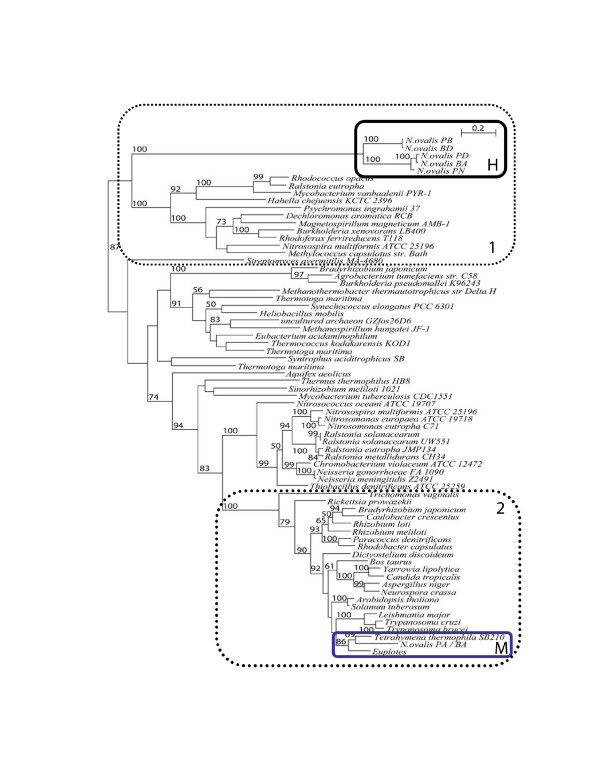
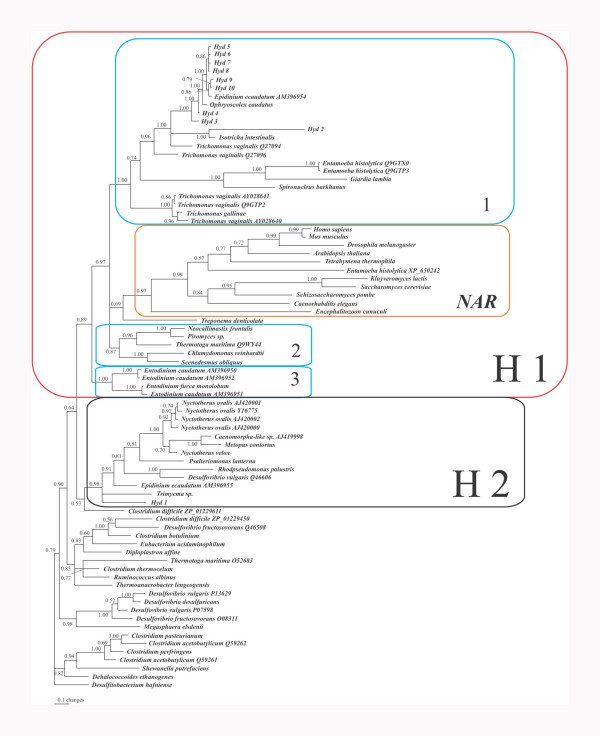
Results: The [FeFe] hydrogenase belongs to a clade of hydrogenases that are different from well-known eukaryotic hydrogenases. The 24 kDa and the 51 kDa modules are most closely related to homologous modules that function in bacterial [NiFe] hydrogenases. Paralogous, mitochondrial 24 kDa and 51 kDa modules function in the mitochondrial complex I in N. ovalis. The different hydrogenase modules have been fused to form a polyprotein that is targeted into the hydrogenosome.
Conclusion: The hydrogenase and their associated modules have most likely been acquired by independent lateral gene transfer from different sources. This scenario for a concerted lateral gene transfer is in agreement with the evolution of the hydrogenosome from a genuine ciliate mitochondrion by evolutionary tinkering.
Figures
References
-
- Müller M. The hydrogenosome. J Gen Microbiol. 1993;139:2879–2889. - PubMed
-
- Roger AJ. Reconstructing Early Events in Eukaryotic Evolution. Am Nat. 1999;154:S146–S163. - PubMed
-
- Hackstein JHP, Akhmanova A, Voncken F, van Hoek A, van Alen T, Boxma B, Moon-van der Staay SY, van der Staay G, Leunissen J, Huynen M, Rosenberg J, Veenhuis M. Hydrogenosomes: convergent adaptations of mitochondria to anaerobic environments. Zoology. 2001;104:290–302. - PubMed
-
- Martin W, Hoffmeister M, Rotte C, Henze K. An overview of endosymbiotic models for the origins of eukaryotes, their ATP-producing organelles (mitochondria and hydrogenosomes), and their heterotrophic lifestyle. Biol Chem. 2001;382:1521–1539. - PubMed
-
- Embley TM, van der Giezen M, Horner DS, Dyal PL, Bell S, Foster PG. Hydrogenosomes, mitochondria and early eukaryotic evolution. IUBMB Life. 2003;55:387–395. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
