

Quantification of ortholog losses in insects and vertebrates
- PMID: 18021399
- PMCID: PMC2258195
- DOI: 10.1186/gb-2007-8-11-r242
Quantification of ortholog losses in insects and vertebrates
Abstract
Background: The increasing number of sequenced insect and vertebrate genomes of variable divergence enables refined comparative analyses to quantify the major modes of animal genome evolution and allows tracing of gene genealogy (orthology) and pinpointing of gene extinctions (losses), which can reveal lineage-specific traits.
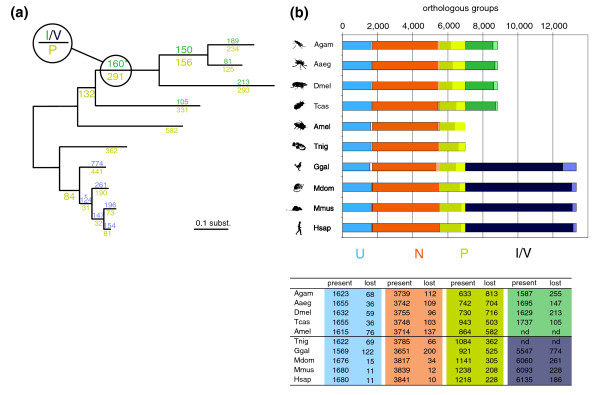
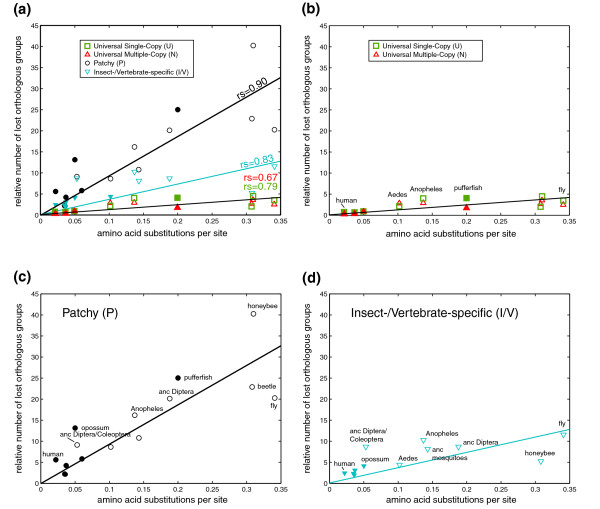
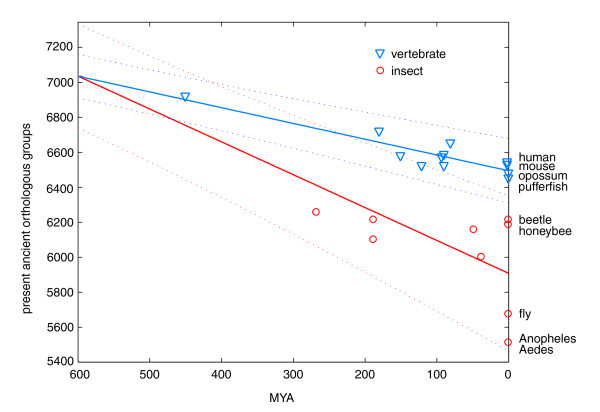
Results: To consistently quantify losses of orthologous groups of genes, we compared the gene repertoires of five vertebrates and five insects, including honeybee and Tribolium beetle, that represent insect orders outside the previously sequenced Diptera. We found hundreds of lost Urbilateria genes in each of the lineages and assessed their phylogenetic origin. The rate of losses correlates well with the species' rates of molecular evolution and radiation times, without distinction between insects and vertebrates, indicating their stochastic nature. Remarkably, this extends to the universal single-copy orthologs, losses of dozens of which have been tolerated in each species. Nevertheless, the propensity for loss differs substantially among genes, where roughly 20% of the orthologs have an 8-fold higher chance of becoming extinct. Extrapolation of our data also suggests that the Urbilateria genome contained more than 7,000 genes.
Conclusion: Our results indicate that the seemingly higher number of observed gene losses in insects can be explained by their two- to three-fold higher evolutionary rate. Despite the profound effect of many losses on cellular machinery, overall, they seem to be guided by neutral evolution.
Figures
References
-
- Zhang J. Evolution by gene duplication: an update. Trends Ecol Evol. 2003;18:292–298. doi: 10.1016/S0169-5347(03)00033-8. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
