

SLIP1, a factor required for activation of histone mRNA translation by the stem-loop binding protein
- PMID: 18025107
- PMCID: PMC2223387
- DOI: 10.1128/MCB.01500-07
SLIP1, a factor required for activation of histone mRNA translation by the stem-loop binding protein
Abstract
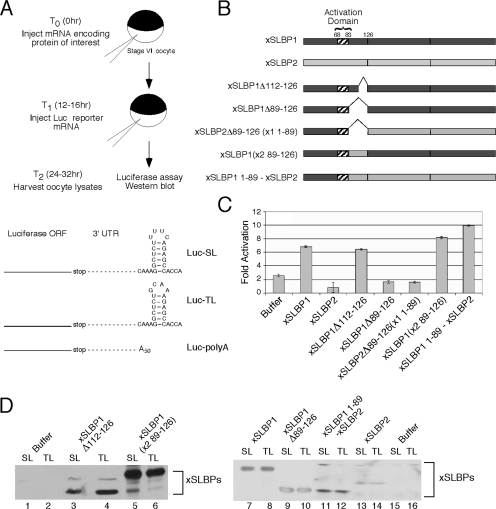
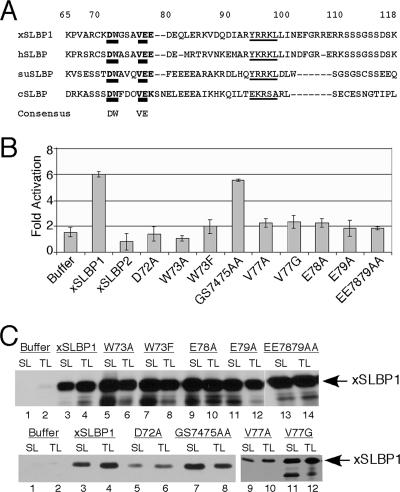
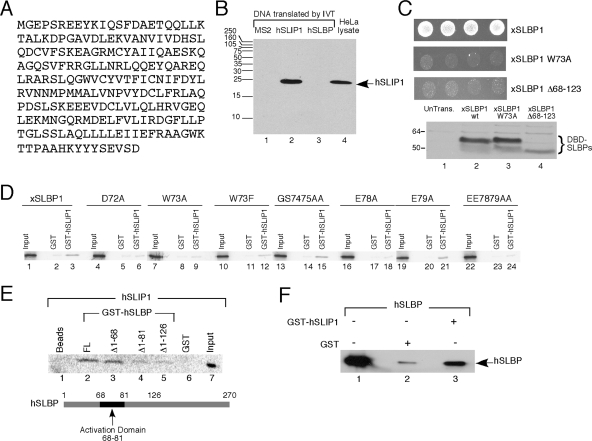
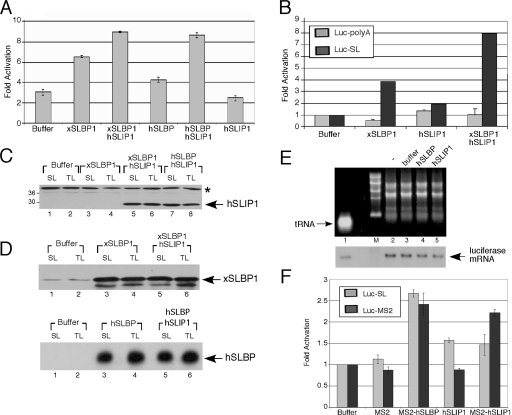
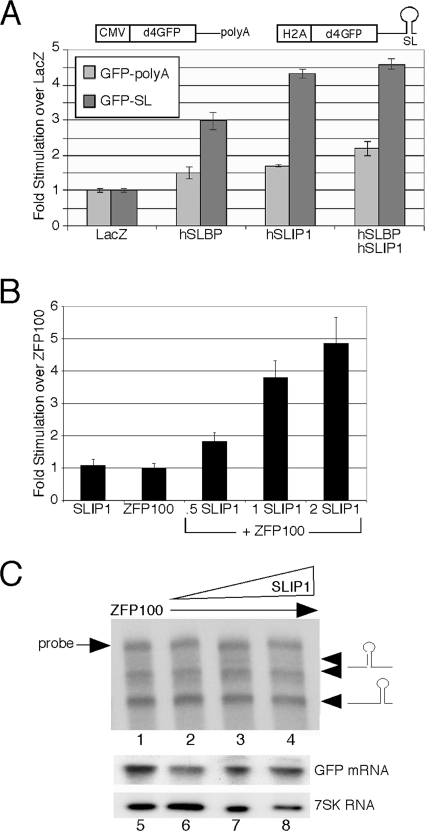
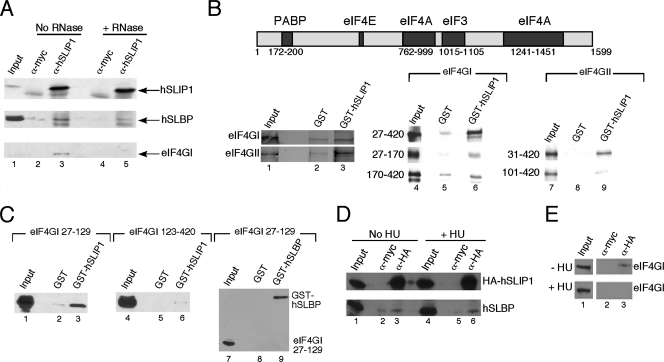
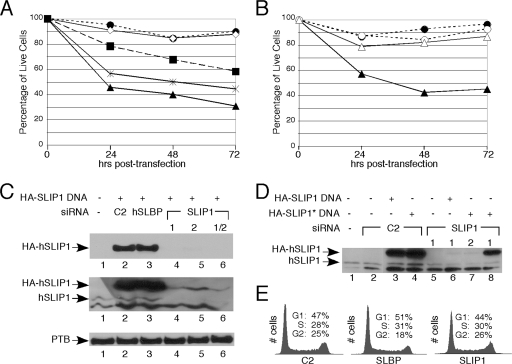
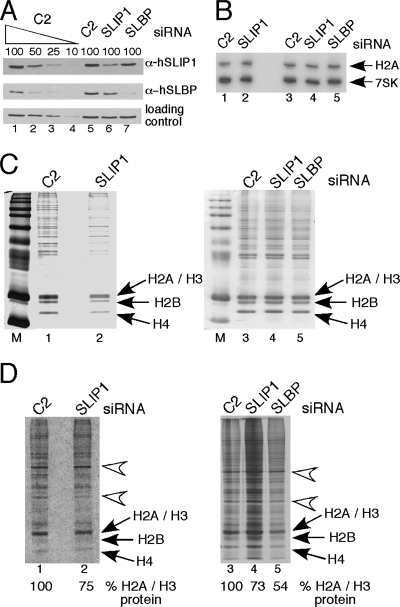
Replication-dependent histone mRNAs are the only eukaryotic cellular mRNAs that are not polyadenylated, ending instead in a conserved stem-loop. The 3' end of histone mRNA is required for histone mRNA translation, as is the stem-loop binding protein (SLBP), which binds the 3' end of histone mRNA. We have identified five conserved residues in a 15-amino-acid region in the amino-terminal portion of SLBP, each of which is required for translation. Using a yeast two-hybrid screen, we identified a novel protein, SLBP-interacting protein 1 (SLIP1), that specifically interacts with this region. Mutations in any of the residues required for translation reduces SLIP1 binding to SLBP. The expression of SLIP1 in Xenopus oocytes together with human SLBP stimulates translation of a reporter mRNA ending in the stem-loop but not a reporter with a poly(A) tail. The expression of SLIP1 in HeLa cells also stimulates the expression of a green fluorescent protein reporter mRNA ending in a stem-loop. RNA interference-mediated downregulation of endogenous SLIP1 reduces the rate of translation of endogenous histone mRNA and also reduces cell viability. SLIP1 may function by bridging the 3' end of the histone mRNA with the 5' end of the mRNA, similar to the mechanism of translation of polyadenylated mRNAs.
Figures
References
-
- Adamson, E. D., and H. R. Woodland. 1977. Changes in the rate of histone synthesis during oocyte maturation and very early development of Xenopus laevis. Dev. Biol. 57136-149. - PubMed
-
- Craig, A. W., A. Haghighat, A. T. Yu, and N. Sonenberg. 1998. Interaction of polyadenylate-binding protein with the eIF4G homologue PAIP enhances translation. Nature 392520-523. - PubMed
-
- Dominski, Z., and W. F. Marzluff. 1999. Formation of the 3′ end of histone mRNA. Gene 2391-14. - PubMed
-
- Gallie, D. R. 1991. The cap and poly(A) tail function synergistically to regulate mRNA translational efficiency. Genes Dev. 52108-2116. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials