

Vesicle formation by self-assembly of membrane-bound matrix proteins into a fluidlike budding domain
- PMID: 18025300
- PMCID: PMC2080896
- DOI: 10.1083/jcb.200705062
Vesicle formation by self-assembly of membrane-bound matrix proteins into a fluidlike budding domain
Abstract
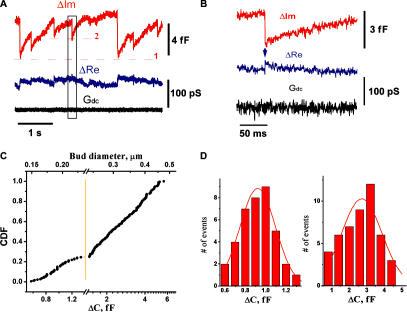
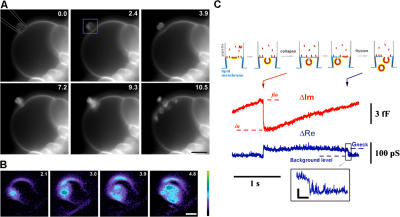
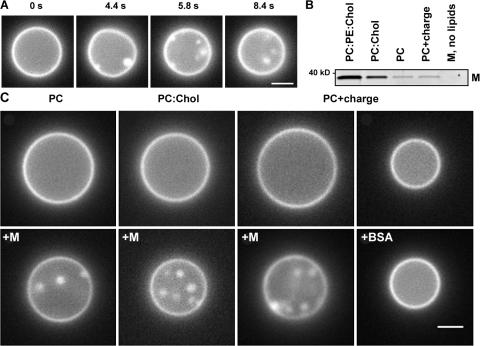
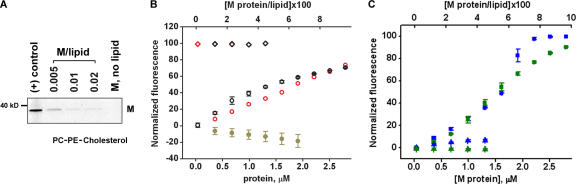
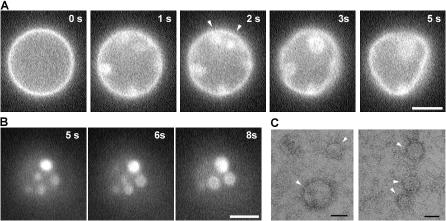
The shape of enveloped viruses depends critically on an internal protein matrix, yet it remains unclear how the matrix proteins control the geometry of the envelope membrane. We found that matrix proteins purified from Newcastle disease virus adsorb on a phospholipid bilayer and condense into fluidlike domains that cause membrane deformation and budding of spherical vesicles, as seen by fluorescent and electron microscopy. Measurements of the electrical admittance of the membrane resolved the gradual growth and rapid closure of a bud followed by its separation to form a free vesicle. The vesicle size distribution, confined by intrinsic curvature of budding domains, but broadened by their merger, matched the virus size distribution. Thus, matrix proteins implement domain-driven mechanism of budding, which suffices to control the shape of these proteolipid vesicles.
Figures
References
-
- Angelova, M.I., and D.S. Dimitrov. 1988. A mechanism of liposome electroformation. Prog. Colloid. Polym. Sci. 76:59–67.
-
- Antonny, B. 2006. Membrane deformation by protein coats. Curr. Opin. Cell Biol. 18:386–394. - PubMed
-
- Basanez, G., J. Zhang, B.N. Chau, G.I. Maksaev, V.A. Frolov, T.A. Brandt, J. Burch, J.M. Hardwick, and J. Zimmerberg. 2001. Pro-apoptotic cleavage products of Bcl-xL form cytochrome c-conducting pores in pure lipid membranes. J. Biol. Chem. 276:31083–31091. - PubMed
-
- Bauer, M., and L. Pelkmans. 2006. A new paradigm for membrane-organizing and shaping scaffolds. FEBS Lett. 580:5559–5564. - PubMed
-
- Bremser, M., W. Nickel, M. Schweikert, M. Ravazzola, M. Amherdt, C.A. Hughes, T.H. Sollner, J.E. Rothman, and F.T. Wieland. 1999. Coupling of coat assembly and vesicle budding to packaging of putative cargo receptors. Cell. 96:495–506. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
