

Retinal adaptation to object motion
- PMID: 18031685
- PMCID: PMC2117331
- DOI: 10.1016/j.neuron.2007.09.030
Retinal adaptation to object motion
Abstract
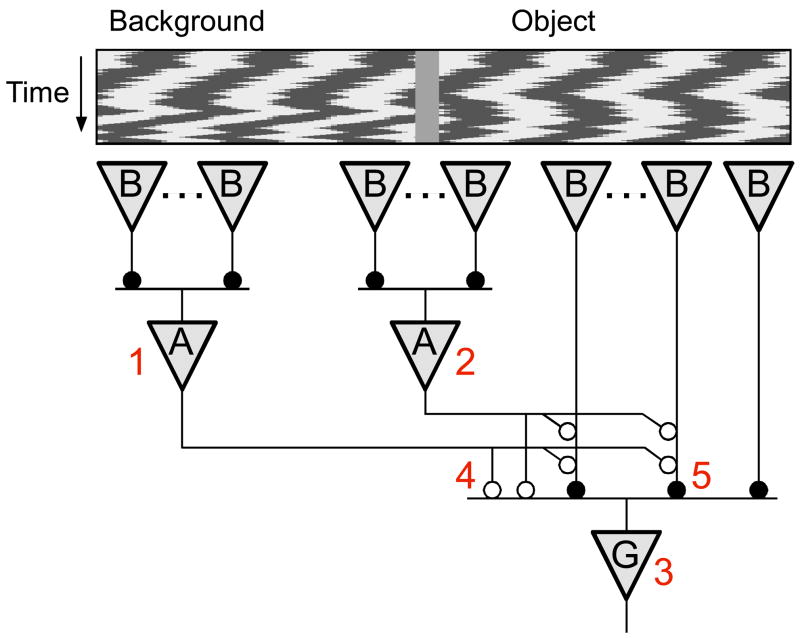
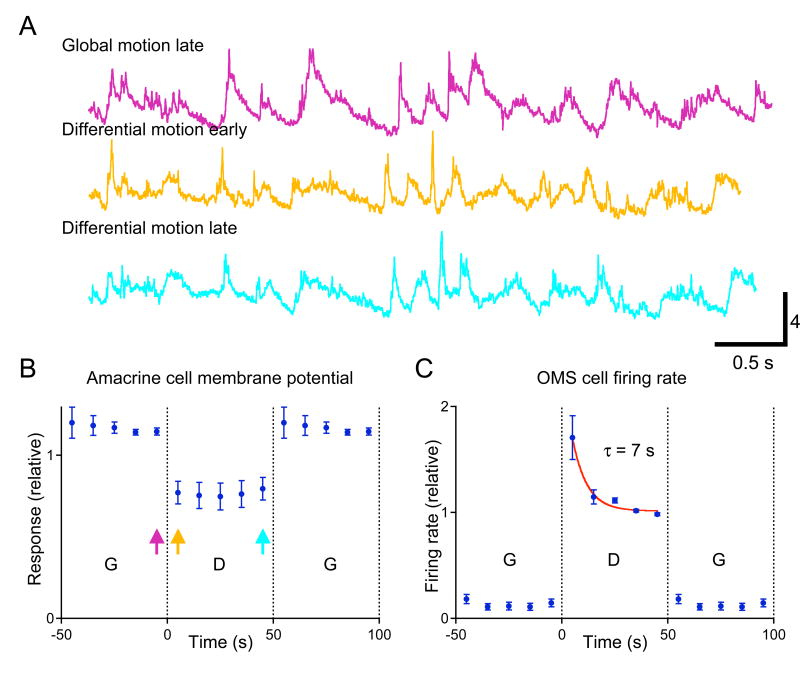
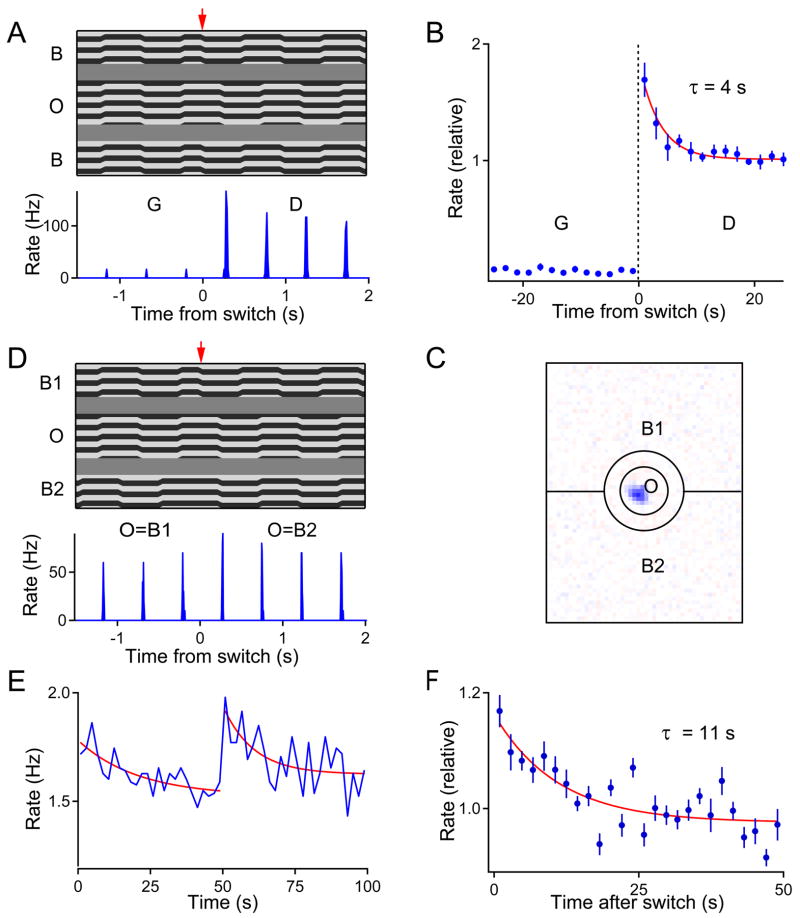
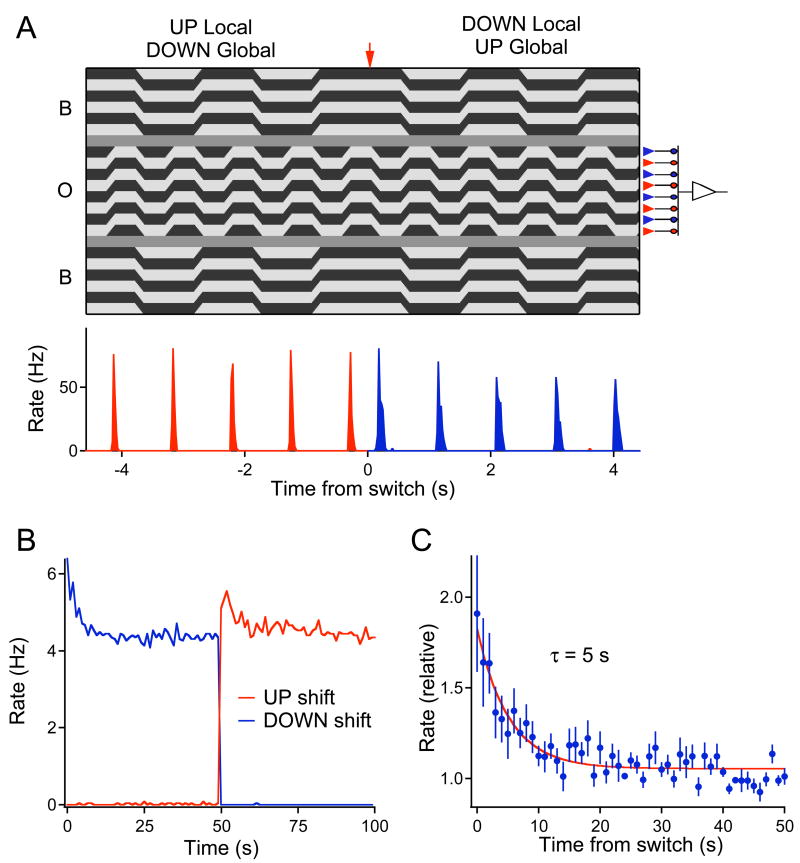
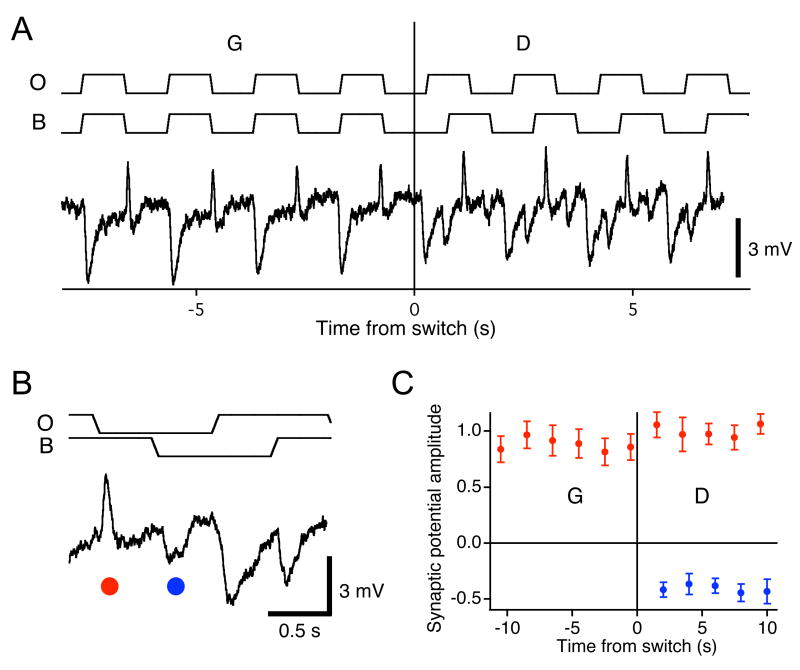
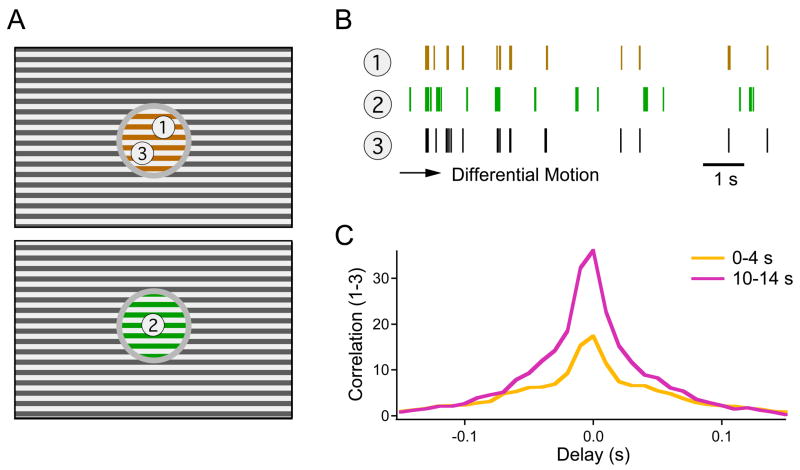
Due to fixational eye movements, the image on the retina is always in motion, even when one views a stationary scene. When an object moves within the scene, the corresponding patch of retina experiences a different motion trajectory than the surrounding region. Certain retinal ganglion cells respond selectively to this condition, when the motion in the cell's receptive field center is different from that in the surround. Here we show that this response is strongest at the very onset of differential motion, followed by gradual adaptation with a time course of several seconds. Different subregions of a ganglion cell's receptive field can adapt independently. The circuitry responsible for differential motion adaptation lies in the inner retina. Several candidate mechanisms were tested, and the adaptation most likely results from synaptic depression at the synapse from bipolar to ganglion cell. Similar circuit mechanisms may act more generally to emphasize novel features of a visual stimulus.
Figures



References
-
- Abrams RA, Christ SE. Motion onset captures attention. Psychol Sci. 2003;14:427–432. - PubMed
-
- Aizenman CD, Huang EJ, Manis PB, Linden DJ. Use-dependent changes in synaptic strength at the Purkinje cell to deep nuclear synapse. Prog Brain Res. 2000;124:257–273. - PubMed
-
- Baccus SA, Meister M. Fast and slow contrast adaptation in retinal circuitry. Neuron. 2002;36:909–919. - PubMed
-
- Barlow HB. A theory about the functional role and synaptic mechanism of visual after-effects. In: Blakemore C, editor. Vision: Coding and Efficiency. Cambridge: Cambridge University Press; 1990. pp. 363–375.
-
- Barlow HB, Hill RM. Evidence for a physiological explanation of the waterfall phenomenon and figural after-effects. Nature. 1963;200:1345–1347. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
