

Lasting changes in a network of interneurons after synapse regeneration and delayed recovery of sensitization
- PMID: 18031937
- PMCID: PMC2198935
- DOI: 10.1016/j.neuroscience.2007.09.061
Lasting changes in a network of interneurons after synapse regeneration and delayed recovery of sensitization
Abstract
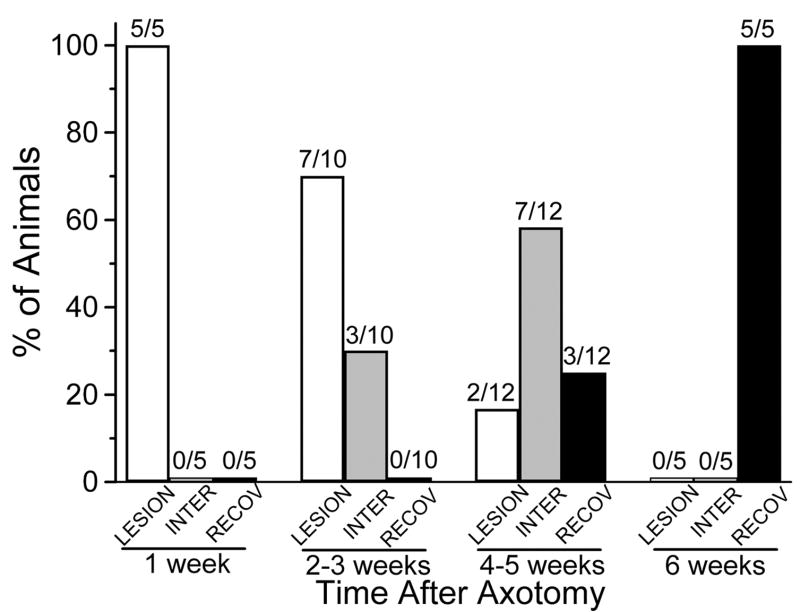
Regeneration of neuronal circuits cannot be successful without restoration of full function, including recovery of behavioral plasticity, which we have found is delayed after regeneration of specific synapses. Experiments were designed to measure neuronal changes that may underlie recovery of function. Sensitization of the leech withdrawal reflex is a non-associative form of learning that depends on the S-interneuron. Cutting an S-cell axon in Faivre's nerve disrupted the capacity for sensitization. The S-cell axon regenerated its electrical synapse with its homologous cell after 3-4 weeks, but the capacity for sensitization was delayed for an additional 2-3 weeks. In the present experiments another form of non-associative conditioning, dishabituation, was also eliminated by S-cell axotomy; it returned following regeneration. Semi-intact preparations were made for behavioral studies, and chains of ganglia with some skin were used for intracellular recording and skin stimulation. In both preparations there was a similar time-course, during 6 weeks, of a lesion-induced decrease and delayed restoration of both S-cell action potential threshold to depolarizing pulses and S-cell firing in response to test stimuli. However, the ability of sensitizing stimuli to decrease S-cell threshold and enhance S-cell activity in response to test stimuli did not fully return after regeneration, indicating that there were lasting changes in the circuit extending beyond the period necessary for full recovery of behavior. Intracellular recordings from the axotomized S-cell revealed a shift in the usual balance of excitatory and inhibitory input, with inhibition enhanced. These results indicate that loss of behavioral plasticity of reflexive shortening following axotomy in the S-cell chain may be related to reduced S-cell activity, and that additional processes underlie full recovery of sensitization of the whole body shortening reflex.
Figures








Similar articles
-
Progressive recovery of learning during regeneration of a single synapse in the medicinal leech.J Comp Neurol. 2003 Feb 24;457(1):67-74. doi: 10.1002/cne.10530. J Comp Neurol. 2003. PMID: 12541325
-
A 3-synapse positive feedback loop regulates the excitability of an interneuron critical for sensitization in the leech.J Neurosci. 2006 Mar 29;26(13):3524-31. doi: 10.1523/JNEUROSCI.3056-05.2006. J Neurosci. 2006. PMID: 16571760 Free PMC article.
-
Neuronal competition for action potential initiation sites in a circuit controlling simple learning.Neuroscience. 2007 Aug 10;148(1):65-81. doi: 10.1016/j.neuroscience.2007.05.046. Epub 2007 Jul 17. Neuroscience. 2007. PMID: 17644266 Free PMC article.
-
Repair and regeneration of functional synaptic connections: cellular and molecular interactions in the leech.Cell Mol Neurobiol. 2005 Mar;25(2):441-50. doi: 10.1007/s10571-005-3152-x. Cell Mol Neurobiol. 2005. PMID: 16047551 Free PMC article. Review.
-
Role of microcircuit structure and input integration in hippocampal interneuron recruitment and plasticity.Neuropharmacology. 2011 Apr;60(5):730-9. doi: 10.1016/j.neuropharm.2010.12.017. Epub 2010 Dec 30. Neuropharmacology. 2011. PMID: 21195097 Review.
References
-
- Blinzinger K, Kreutzberg G. Displacement of synaptic terminals from regenerating motoneurons by microglial cells. Z Zellforsch Mikrosk Anat. 1968;85:145–157. - PubMed
-
- Bristol AS, Sutton MA, Carew TJ. Neural circuit of tail-elicited siphon withdrawal in Aplysia. I. Differential lateralization of sensitization and dishabituation. J Neurophysiol. 2004;91:666–677. - PubMed
-
- Burrell BD, Sahley CL, Muller KJ. Differential effects of serotonin enhance activity of an electrically coupled neural network. J Neurophysiol. 2002;87:2889–2895. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
