

NR2B- and NR2D-containing synaptic NMDA receptors in developing rat substantia nigra pars compacta dopaminergic neurones
- PMID: 18033813
- PMCID: PMC2375608
- DOI: 10.1113/jphysiol.2007.144618
NR2B- and NR2D-containing synaptic NMDA receptors in developing rat substantia nigra pars compacta dopaminergic neurones
Abstract
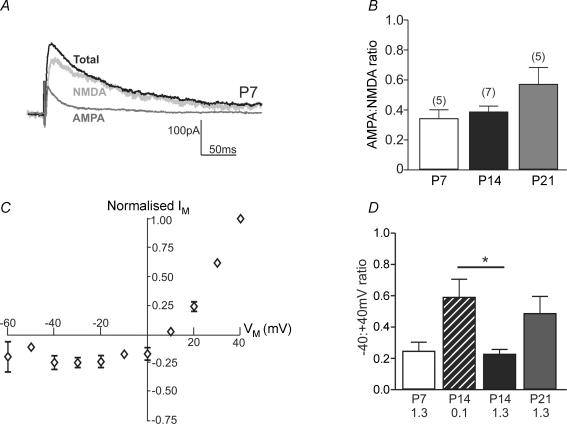
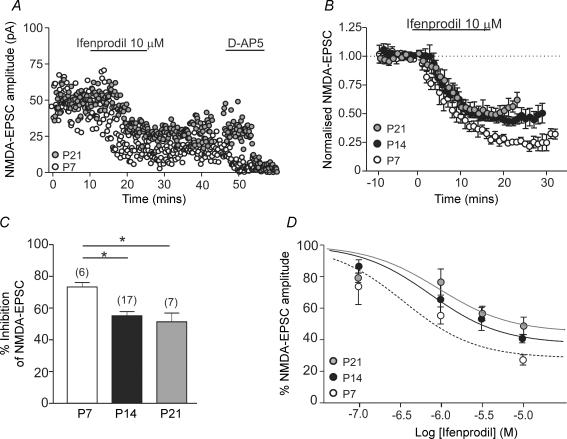
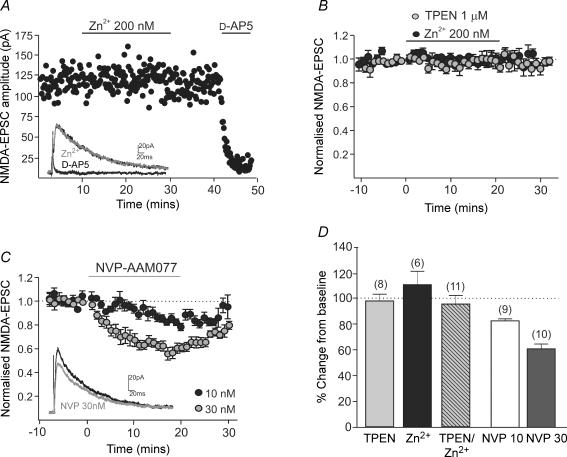
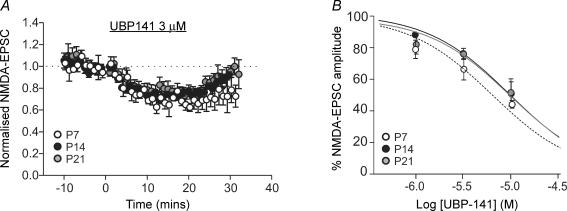
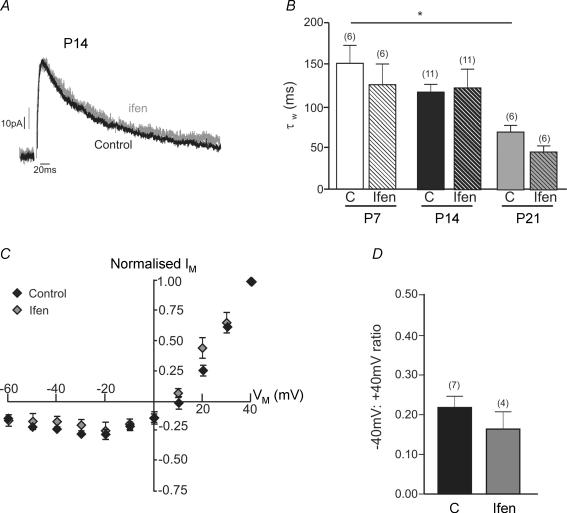
NMDA receptors are present at glutamatergic synapses throughout the brain, and are important for the development and plasticity of neural circuits. Their subunit composition is developmentally regulated. We have investigated the developmental profile of functional synaptic NMDA receptor subunits in dopaminergic neurones of the substantia nigra pars compacta (SNc). In SNc dopaminergic neurones from rats aged postnatal day (P)7, ifenprodil inhibited NMDA-EPSCs with an estimated IC(50) of 0.36 microm and a maximum inhibition of 73.5 +/- 2.7% (10 microm), consistent with a substantial population of NR1/NR2B-containing diheteromeric receptors. UBP141, a novel NR2D-preferring antagonist, inhibited NMDA-EPSCs with an estimated IC(50) of 6.2 microm. During postnatal development, the maximum inhibitory effect of 10 microm ifenprodil significantly decreased. However, NMDA-EPSCs were not inhibited by Zn(2+) (200 nM) or potentiated by the Zn(2+) chelator TPEN (1 microm), and the effect of UBP141 did not increase during development, indicating that NR2B subunits are not replaced with diheteromeric NR2A or NR2D subunits. The time course of the decay of NMDA-EPSCs was not significantly changed in ifenprodil at any age tested. Together, these data suggest that diheteromeric NR1/NR2A or NR1/NR2D receptors do not account for the ifenprodil-resistant component of the NMDA-EPSC. We propose that NR1/NR2B/NR2D triheteromers form a significant fraction of synaptic NMDA receptors during postnatal development. This is the first report of data suggesting NR2D-containing triheteromeric NMDA receptors at a brain synapse.
Figures
Comment in
-
2B or 2B and 2D? - that is the question.J Physiol. 2008 Feb 1;586(3):693. doi: 10.1113/jphysiol.2007.148924. J Physiol. 2008. PMID: 18245207 Free PMC article. No abstract available.
References
-
- Albers DS, Weiss SW, Ladarola MJ, Standaert DG. Immunohistochemical localization of N-methyl-D-aspartate and α-amino-3-hydroxy-5-methyl-4-isoxazolepropionate receptor subunits in the substantia nigra pars compacta of the rat. Neuroscience. 1999;89:209–220. - PubMed
-
- Auberson YP, Allgeier H, Bischoff S, Lingenhoehl K, Moretti R, Schmutz M. 5-Phosphonomethylquinoxalinediones as competitive NMDA receptor antagonists with a preference for the human 1A/2A, rather than 1A/2B receptor composition. Bioorg Med Chem Lett. 2002;12:1099–1102. - PubMed
-
- Blanchet PJ, Konitsiotis S, Whittemore ER, Zhou ZL, Woodward RM, Chase TN. Differing effects of N-methyl-D-aspartate receptor subtype selective antagonists on dyskinesias in levodopa-treated 1-methyl-4-phenyl-tetrahydropyridine monkeys. J Pharmacol Exp Ther. 1999;290:1034–1040. - PubMed
-
- Blandini F, Nappi G, Tassorelli C, Martignoni E. Functional changes of the basal ganglia circuitry in Parkinson's disease. Prog Neurobiol. 2000;62:63–88. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- MH060252-05A2/MH/NIMH NIH HHS/United States
- R01 MH060252/MH/NIMH NIH HHS/United States
- G0601812/MRC_/Medical Research Council/United Kingdom
- BB/D015782/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/D015286/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Miscellaneous