

Genome-wide transcriptional analysis of grapevine berry ripening reveals a set of genes similarly modulated during three seasons and the occurrence of an oxidative burst at vèraison
- PMID: 18034875
- PMCID: PMC2228314
- DOI: 10.1186/1471-2164-8-428
Genome-wide transcriptional analysis of grapevine berry ripening reveals a set of genes similarly modulated during three seasons and the occurrence of an oxidative burst at vèraison
Abstract
Background: Grapevine (Vitis species) is among the most important fruit crops in terms of cultivated area and economic impact. Despite this relevance, little is known about the transcriptional changes and the regulatory circuits underlying the biochemical and physical changes occurring during berry development.
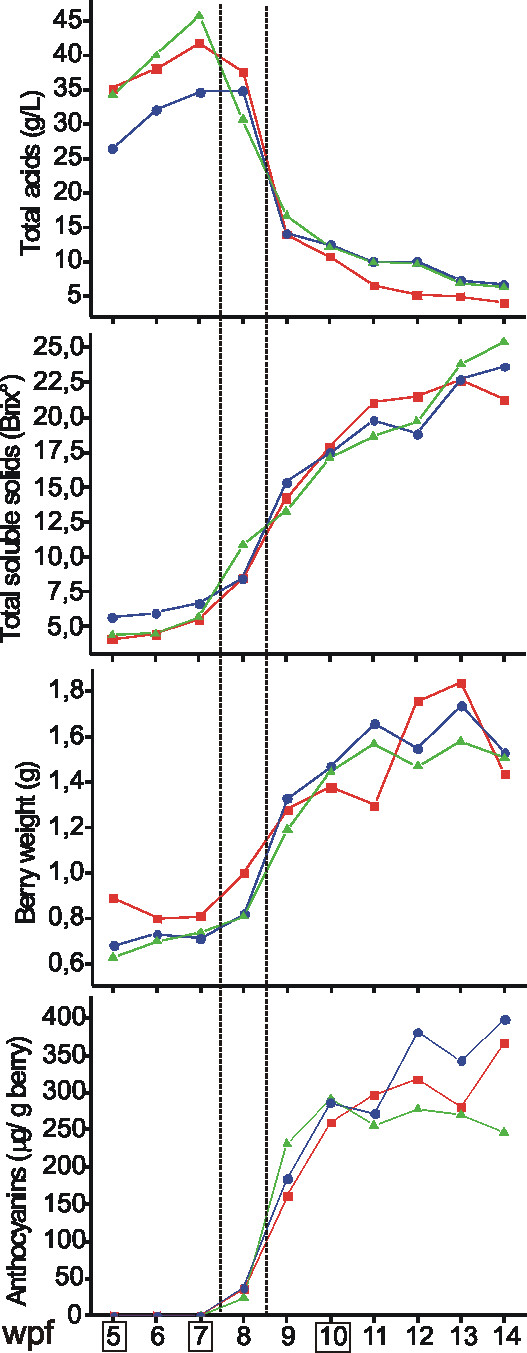
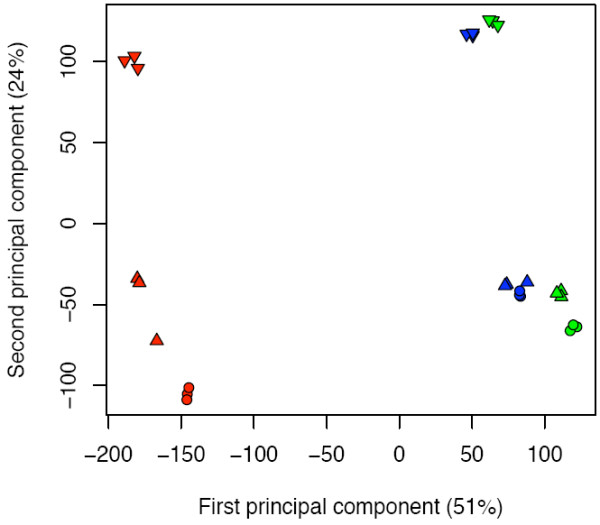
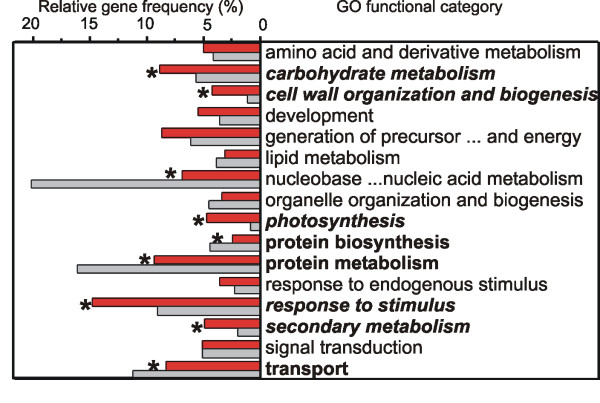
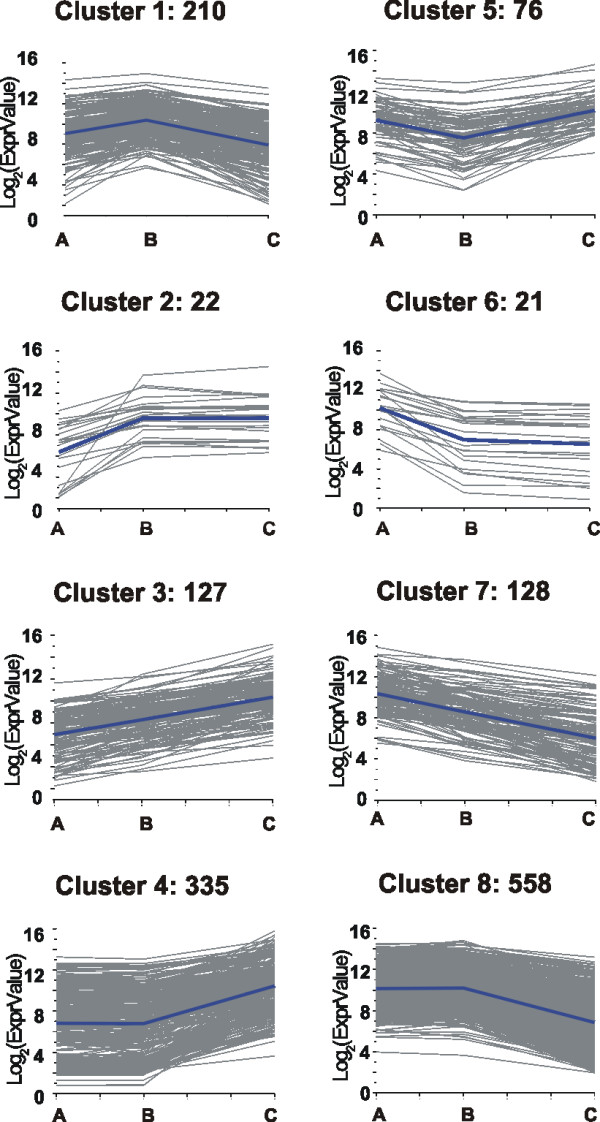
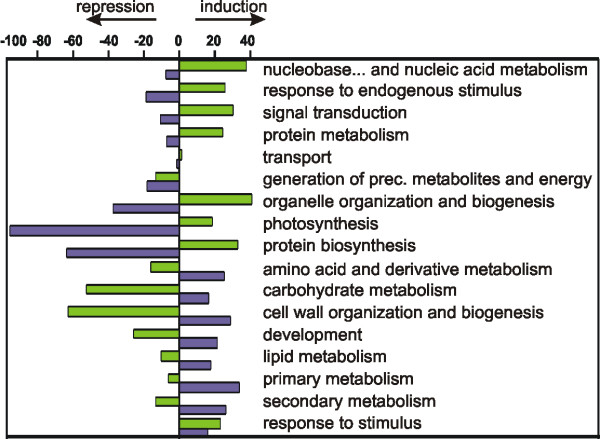
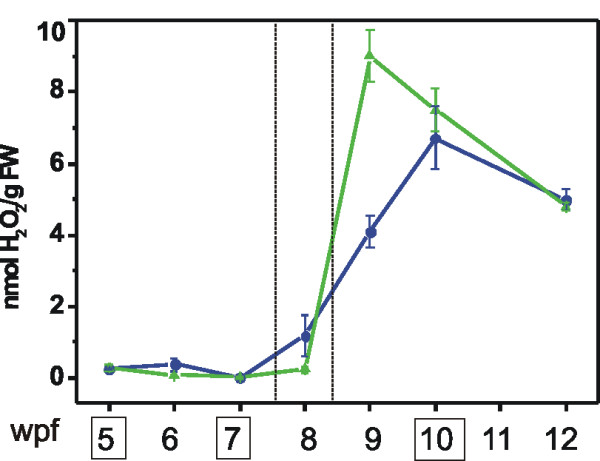
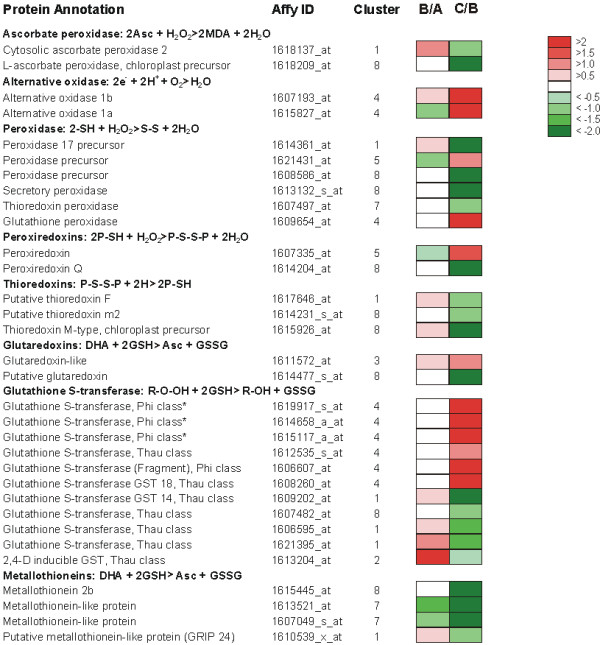
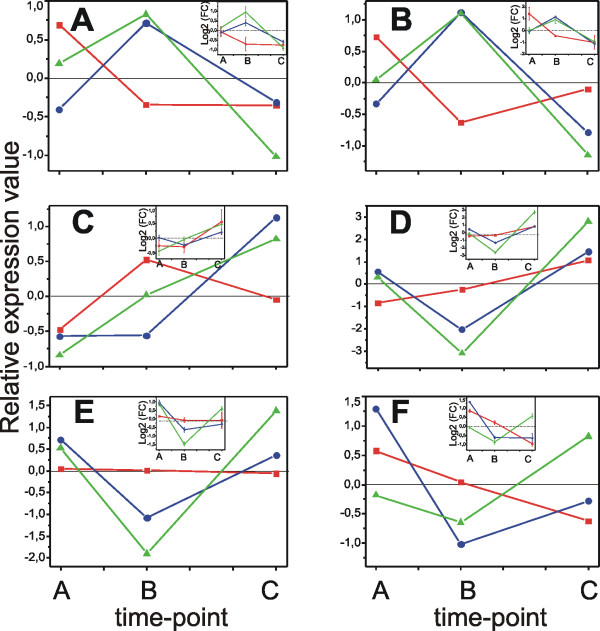
Results: Fruit ripening in the non-climacteric crop species Vitis vinifera L. has been investigated at the transcriptional level by the use of the Affymetrix Vitis GeneChip which contains approximately 14,500 unigenes. Gene expression data obtained from berries sampled before and after véraison in three growing years, were analyzed to identify genes specifically involved in fruit ripening and to investigate seasonal influences on the process. From these analyses a core set of 1477 genes was found which was similarly modulated in all seasons. We were able to separate ripening specific isoforms within gene families and to identify ripening related genes which appeared strongly regulated also by the seasonal weather conditions. Transcripts annotation by Gene Ontology vocabulary revealed five overrepresented functional categories of which cell wall organization and biogenesis, carbohydrate and secondary metabolisms and stress response were specifically induced during the ripening phase, while photosynthesis was strongly repressed. About 19% of the core gene set was characterized by genes involved in regulatory processes, such as transcription factors and transcripts related to hormonal metabolism and signal transduction. Auxin, ethylene and light emerged as the main stimuli influencing berry development. In addition, an oxidative burst, previously not detected in grapevine, characterized by rapid accumulation of H2O2 starting from véraison and by the modulation of many ROS scavenging enzymes, was observed.
Conclusion: The time-course gene expression analysis of grapevine berry development has identified the occurrence of two well distinct phases along the process. The pre-véraison phase represents a reprogramming stage of the cellular metabolism, characterized by the expression of numerous genes involved in hormonal signalling and transcriptional regulation. The post-véraison phase is characterized by the onset of a ripening-specialized metabolism responsible for the phenotypic traits of the ripe berry. Between the two phases, at véraison, an oxidative burst and the concurrent modulation of the anti-oxidative enzymatic network was observed. The large number of regulatory genes we have identified represents a powerful new resource for dissecting the mechanisms of fruit ripening control in non-climacteric plants.
Figures
References
-
- IASMA Genomics http://genomics.research.iasma.it
-
- NCBI dbest http://www.ncbi.nlm.nih.gov/dbEST/
-
- Coombe BG, Mccarthy MG. Dynamics of Grape Berry Growth and Physiology of Ripening. Aust J Grape Wine Res. 2000;6:131–135.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
