

Restriction endonuclease inhibitor IPI* of bacteriophage T4: a novel structure for a dedicated target
- PMID: 18037438
- PMCID: PMC2255585
- DOI: 10.1016/j.jmb.2007.10.064
Restriction endonuclease inhibitor IPI* of bacteriophage T4: a novel structure for a dedicated target
Abstract
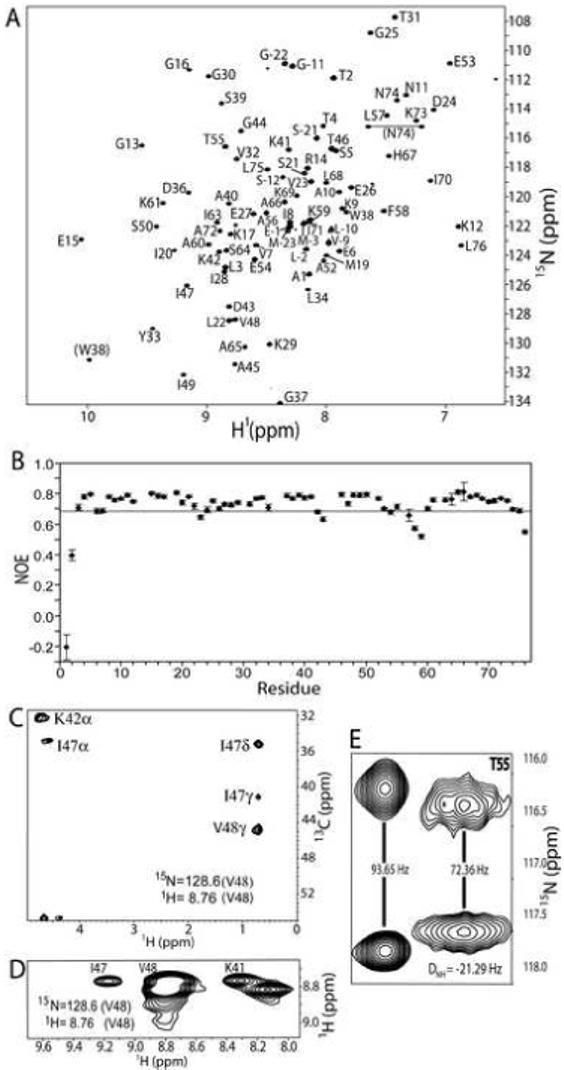
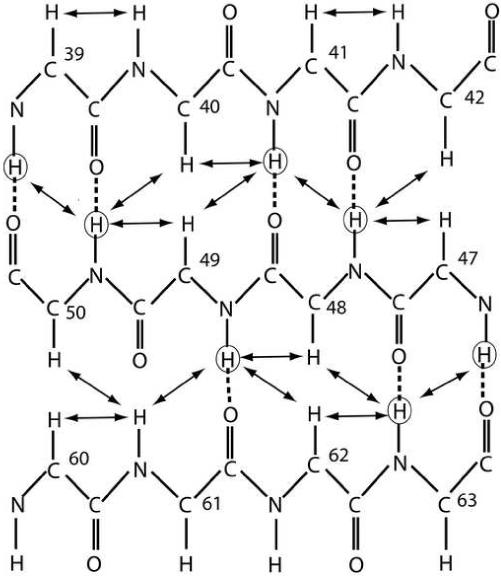
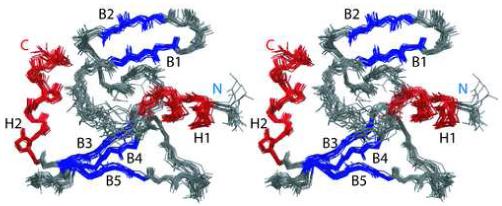
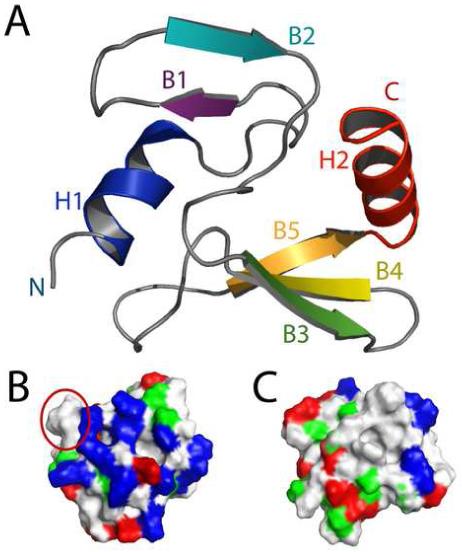
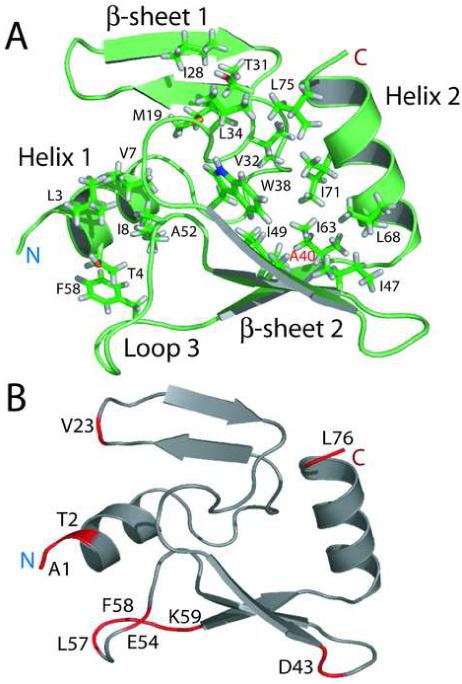
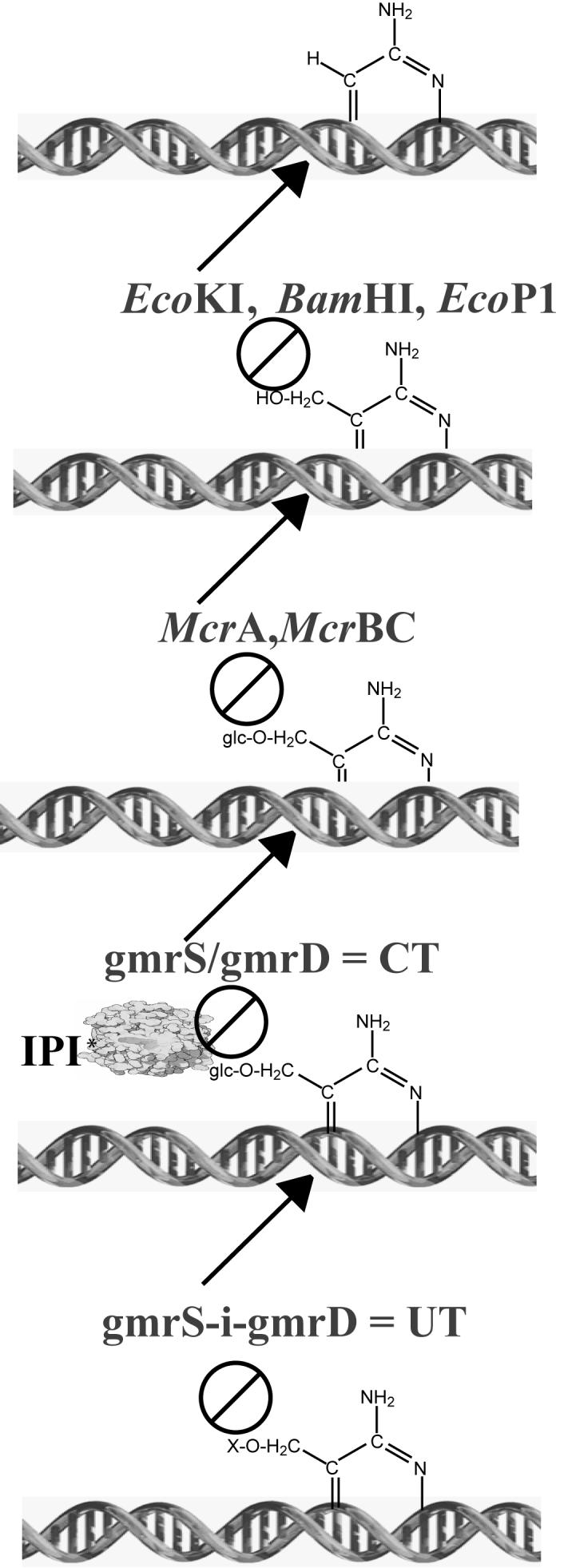
Phage T4 protects its DNA from the two-gene-encoded gmrS/gmrD (glucose-modified hydroxymethylcytosine restriction endonuclease) CT of pathogenic Escherichia coli, CT596, by injecting several hundred copies of the 76-amino-acid-residue nuclease inhibitor, IPI*, into the infected host. Here, the three-dimensional solution structure of mature IPI* is reported as determined by nuclear magnetic resonance techniques using 1290 experimental nuclear Overhauser effect and dipolar coupling constraints ( approximately 17 constraints per residue). Close examination of this oblate-shaped protein structure reveals a novel fold consisting of two small beta-sheets (beta1: B1 and B2; beta2: B3-B5) flanked at the N- and C-termini by alpha-helices (H1 and H2). Such a fold is very compact in shape and allows ejection of IPI* through the narrow 30-A portal and tail tube apertures of the virion without unfolding. Structural and dynamic measurements identify an exposed hydrophobic knob that is a putative gmrS/gmrD-binding site. A single gene from the uropathogenic E. coli UT189, which codes for a gmrS/gmrD-like UT fusion enzyme (with approximately 90% identity to the heterodimeric CT enzyme), has evolved IPI* inhibitor immunity. Analysis of the gmrS/gmrD restriction endonuclease enzyme family and its IPI* family phage antagonists reveals an evolutionary pathway that has elaborated a surprisingly diverse and specifically fitted set of coevolving attack and defense structures.
Figures

References
-
- Abremski K, Black LW. The function of bacteriophage T4 internal protein I in a restrictive strain of Escherichia coli. Virology. 1979;97(2):439–47. - PubMed
-
- Mullaney JM, Black LW. Capsid targeting sequence targets foreign proteins into bacteriophage T4 and permits proteolytic processing. J Mol Biol. 1996;261(3):372–85. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
