

MUC1 oncoprotein activates the IkappaB kinase beta complex and constitutive NF-kappaB signalling
- PMID: 18037881
- PMCID: PMC4209910
- DOI: 10.1038/ncb1661
MUC1 oncoprotein activates the IkappaB kinase beta complex and constitutive NF-kappaB signalling
Abstract
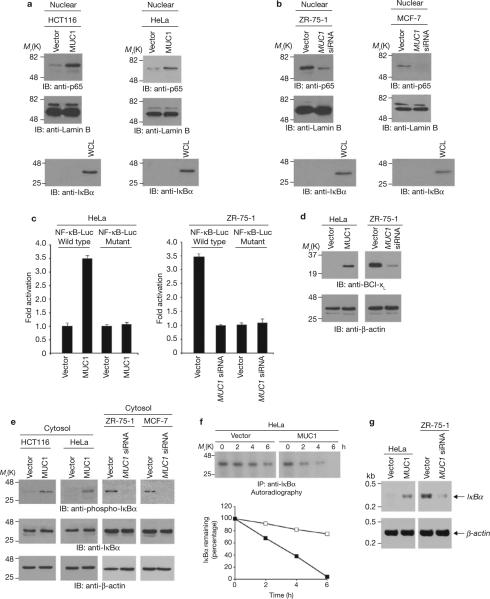
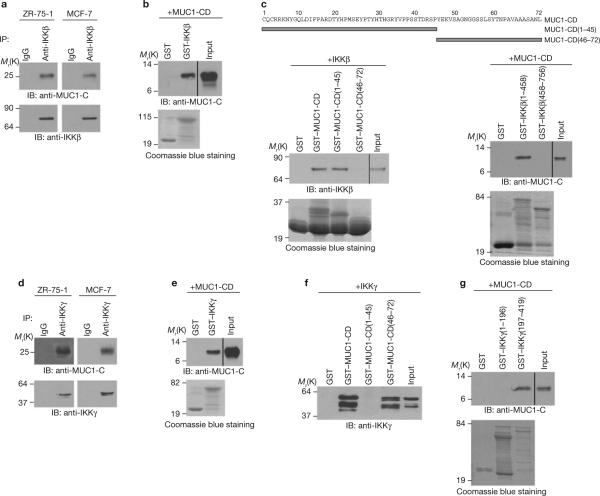
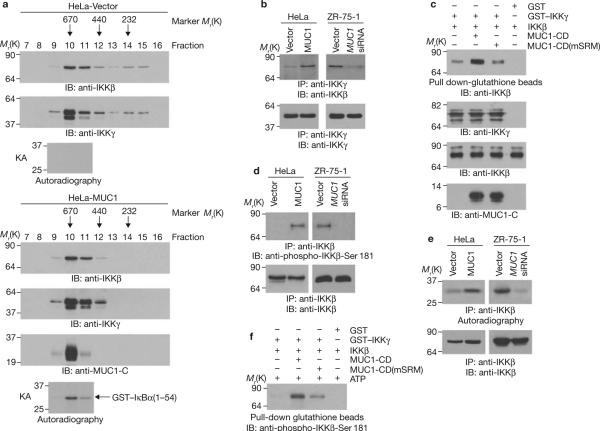
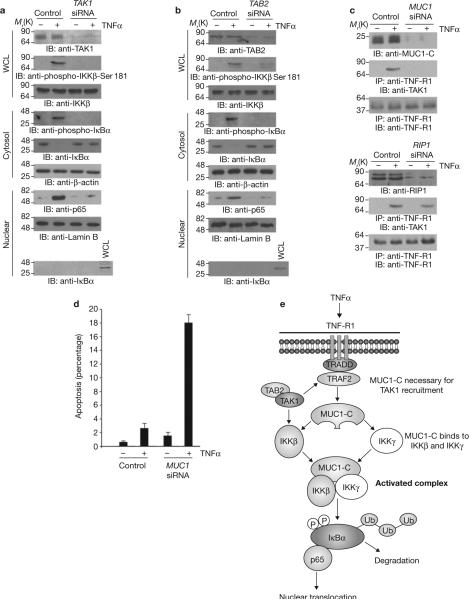
Nuclear factor-kappaB (NF-kappaB) is constitutively activated in diverse human malignancies by mechanisms that are not understood. The MUC1 oncoprotein is aberrantly overexpressed by most human carcinomas and, similarly to NF-kappaB, blocks apoptosis and induces transformation. This study demonstrates that overexpression of MUC1 in human carcinoma cells is associated with constitutive activation of NF-kappaB p65. We show that MUC1 interacts with the high-molecular-weight IkappaB kinase (IKK) complex in vivo and that the MUC1 cytoplasmic domain binds directly to IKKbeta and IKKgamma. Interaction of MUC1 with both IKKbeta and IKKgamma is necessary for IKKbeta activation, resulting in phosphorylation and degradation of IkappaBalpha. Studies in non-malignant epithelial cells show that MUC1 is recruited to the TNF-R1 complex and interacts with IKKbeta-IKKgamma in response to TNFalpha stimulation. TNFalpha-induced recruitment of MUC1 is dependent on TRADD and TRAF2, but not the death-domain kinase RIP1. In addition, MUC1-mediated activation of IKKbeta is dependent on TAK1 and TAB2. These findings indicate that MUC1 is important for physiological activation of IKKbeta and that overexpression of MUC1, as found in human cancers, confers sustained induction of the IKKbeta-NF-kappaB p65 pathway.
Figures

References
-
- Yamamoto Y, Gaynor R. IκB kinases: key regulators of the NF-κB pathway. Trends Biochem. Sci. 2003;29:72–79. - PubMed
-
- Hayden MS, Ghosh S. Signaling to NF-κB. Genes Dev. 2004;18:2195–2224. - PubMed
-
- Wei X, Xu H, Kufe D. Human MUC1 oncoprotein regulates p53-responsive gene transcription in the genotoxic stress response. Cancer Cell. 2005;7:167–178. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
