

Identification of heme as the ligand for the orphan nuclear receptors REV-ERBalpha and REV-ERBbeta
- PMID: 18037887
- PMCID: PMC2743565
- DOI: 10.1038/nsmb1344
Identification of heme as the ligand for the orphan nuclear receptors REV-ERBalpha and REV-ERBbeta
Abstract
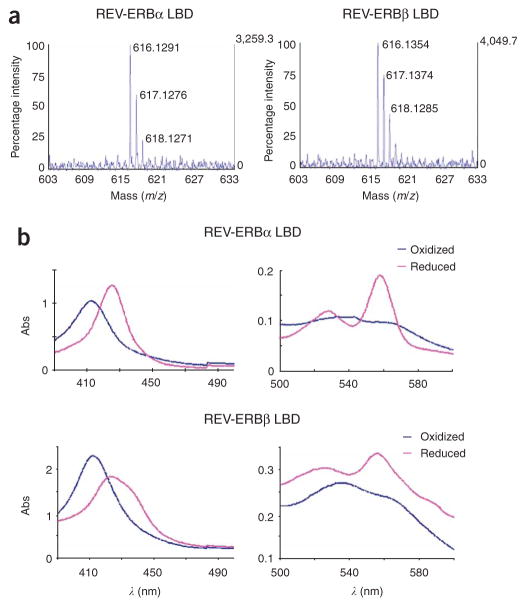
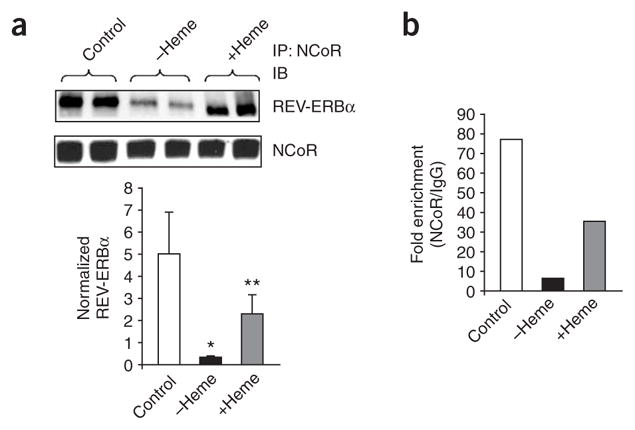
The nuclear receptors REV-ERBalpha (encoded by NR1D1) and REV-ERBbeta (NR1D2) have remained orphans owing to the lack of identified physiological ligands. Here we show that heme is a physiological ligand of both receptors. Heme associates with the ligand-binding domains of the REV-ERB receptors with a 1:1 stoichiometry and enhances the thermal stability of the proteins. Results from experiments of heme depletion in mammalian cells indicate that heme binding to REV-ERB causes the recruitment of the co-repressor NCoR, leading to repression of target genes including BMAL1 (official symbol ARNTL), an essential component of the circadian oscillator. Heme extends the known types of ligands used by the human nuclear receptor family beyond the endocrine hormones and dietary lipids described so far. Our results further indicate that heme regulation of REV-ERBs may link the control of metabolism and the mammalian clock.
Figures



References
-
- Miyajima N, et al. Two erbA homologs encoding proteins with different T3 binding capacities are transcribed from opposite DNA strands of the same genetic locus. Cell. 1989;57:31–39. - PubMed
-
- Bonnelye E, et al. Rev-erbβ, a new member of the nuclear receptor superfamily, is expressed in the nervous system during chicken development. Cell Growth Differ. 1994;5:1357–1365. - PubMed
-
- Dumas B, et al. A new orphan member of the nuclear hormone receptor superfamily closely related to Rev-Erb. Mol Endocrinol. 1994;8:996–1005. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
