

Multivalent engagement of chromatin modifications by linked binding modules
- PMID: 18037899
- PMCID: PMC4690530
- DOI: 10.1038/nrm2298
Multivalent engagement of chromatin modifications by linked binding modules
Abstract
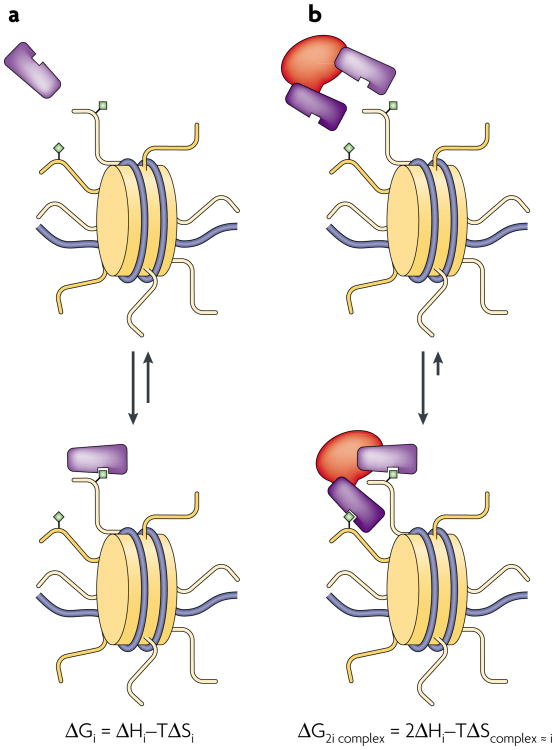
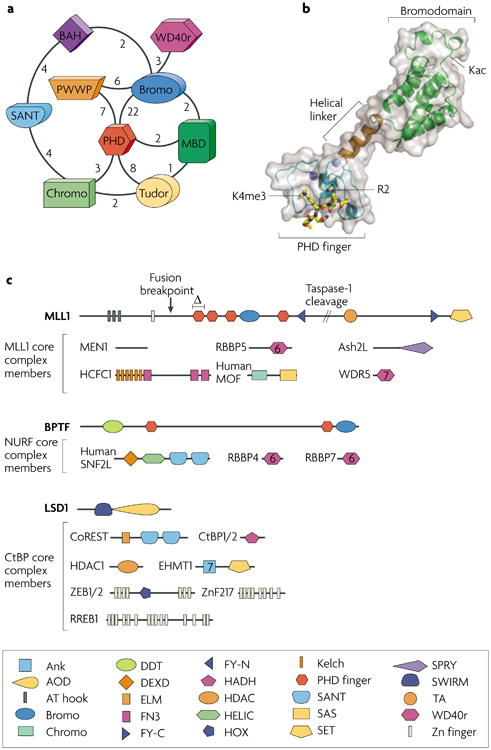
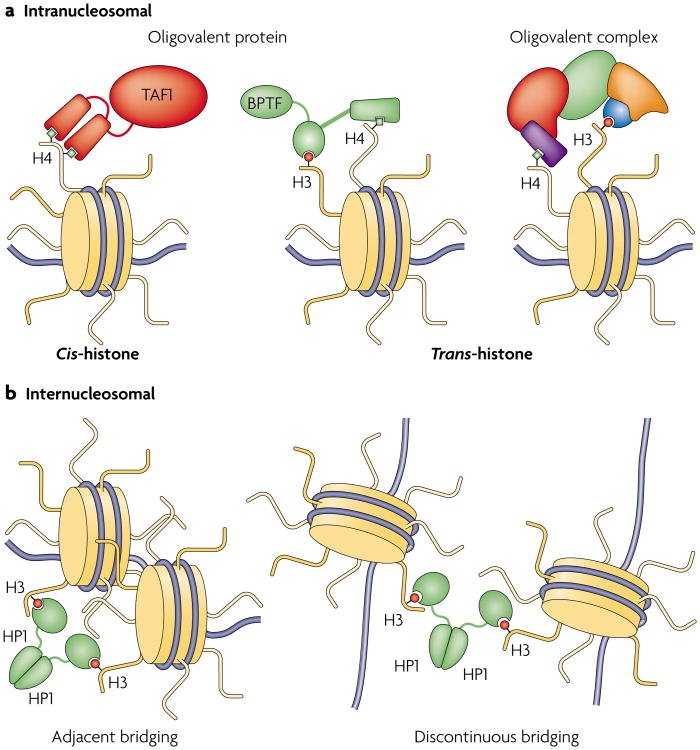
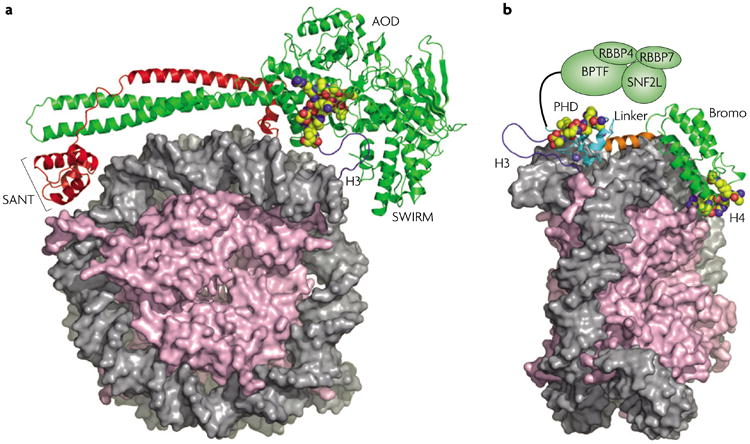
Various chemical modifications on histones and regions of associated DNA play crucial roles in genome management by binding specific factors that, in turn, serve to alter the structural properties of chromatin. These so-called effector proteins have typically been studied with the biochemist's paring knife--the capacity to recognize specific chromatin modifications has been mapped to an increasing number of domains that frequently appear in the nuclear subset of the proteome, often present in large, multisubunit complexes that bristle with modification-dependent binding potential. We propose that multivalent interactions on a single histone tail and beyond may have a significant, if not dominant, role in chromatin transactions.
Figures
References
-
- Cosgrove MS. Histone proteomics and the epigenetic regulation of nucleosome mobility. Expert Rev Proteomics. 2007;4:465–478. A recent overview of known histone PTMs as identified by mass spectrometry It focuses on direct nucleosome-intrinsic effects of histone modifications and represents a nice catalogue of marks. - PubMed
-
- Clarkson MJ, Wells JR, Gibson F, Saint R, Tremethick DJ. Regions of variant histone His2AvD required for Drosophila development. Nature. 1999;399:694–697. - PubMed
-
- Bernstein E, Hake SB. The nucleosome: a little variation goes a long way. Biochem Cell Biol. 2006;84:505–517. - PubMed
-
- Henikoff S, Ahmad K. Assembly of variant histones into chromatin. Annu Rev Cell Dev Biol. 2005;21:133–153. A definitive review on histone variants that describes the functional associations of variants with various stages of chromatin metabolism. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
