

Phylogenetic footprinting analysis in the upstream regulatory regions of the Drosophila enhancer of split genes
- PMID: 18039873
- PMCID: PMC2147979
- DOI: 10.1534/genetics.107.070425
Phylogenetic footprinting analysis in the upstream regulatory regions of the Drosophila enhancer of split genes
Abstract
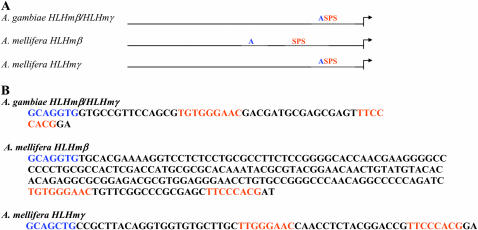
During Drosophila development Suppressor of Hairless [Su(H)]-dependent Notch activation upregulates transcription of the Enhancer of split-Complex [E(spl)-C] genes. Drosophila melanogaster E(spl) genes share common transcription regulators including binding sites for Su(H), proneural, and E(spl) basic-helix-loop-helix (bHLH) proteins. However, the expression patterns of E(spl) genes during development suggest that additional factors are involved. To better understand regulators responsible for these expression patterns, recently available sequence and annotation data for multiple Drosophila genomes were used to compare the E(spl) upstream regulatory regions from more than nine Drosophila species. The mgamma and mbeta regulatory regions are the most conserved of the bHLH genes. Fine analysis of Su(H) sites showed that high-affinity Su(H) paired sites and the Su(H) paired site plus proneural site (SPS + A) architecture are completely conserved in a subset of Drosophila E(spl) genes. The SPS + A module is also present in the upstream regulatory regions of the more ancient mosquito and honeybee E(spl) bHLH genes. Additional transcription factor binding sites were identified upstream of the E(spl) genes and compared between species of Drosophila. Conserved sites provide new understandings about E(spl) regulation during development. Conserved novel sequences found upstream of multiple E(spl) genes may play a role in the expression of these genes.
Figures






References
-
- Artavanis-Tsakonas, S., M. D. Rand and R. J. Lake, 1999. Notch signaling: cell fate control and signal integration in development. Science 284: 770–776. - PubMed
-
- Axelson, H., 2004. a Notch signaling and cancer: emerging complexity. Semin. Cancer Biol. 14: 317–319. - PubMed
-
- Axelson, H., 2004. b The Notch signaling cascade in neuroblastoma: role of the basic helix-loop-helix proteins HASH-1 and HES-1. Cancer Lett. 204: 171–178. - PubMed
-
- Bailey, A. M., and J. W. Posakony, 1995. Suppressor of hairless directly activates transcription of enhancer of split complex genes in response to Notch receptor activity. Genes Dev. 9: 2609–2622. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
