

In vivo visualization of type II plasmid segregation: bacterial actin filaments pushing plasmids
- PMID: 18039937
- PMCID: PMC2099209
- DOI: 10.1083/jcb.200708206
In vivo visualization of type II plasmid segregation: bacterial actin filaments pushing plasmids
Abstract
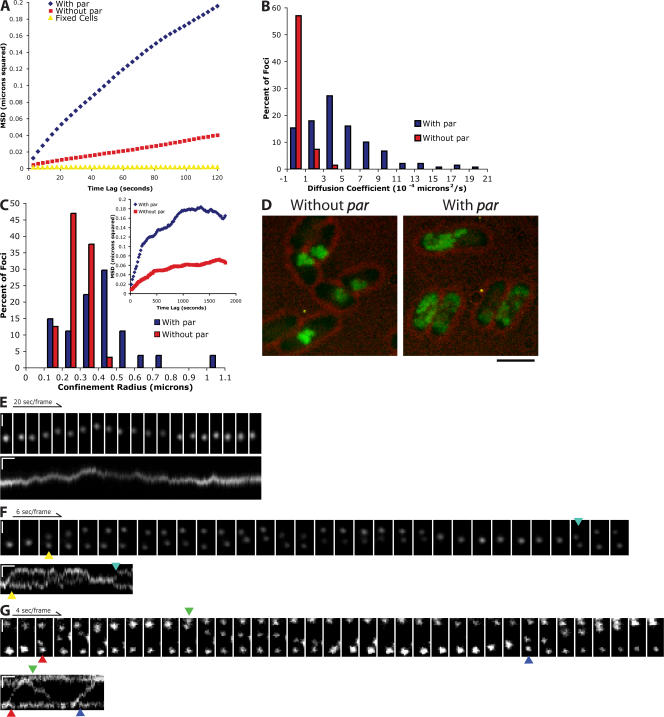
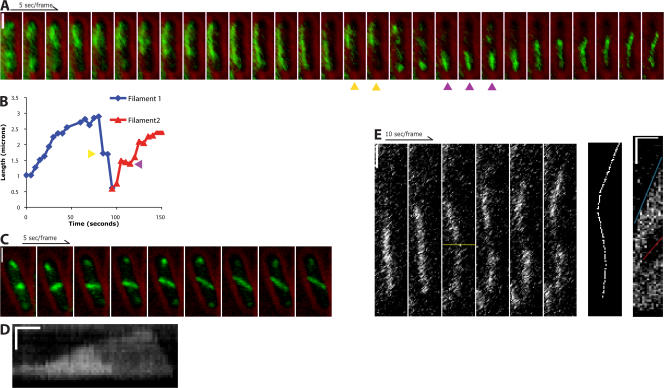
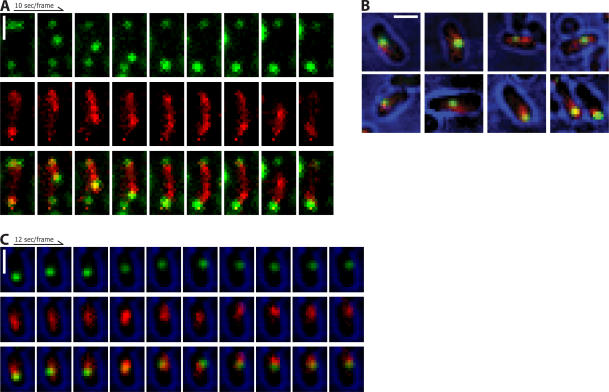
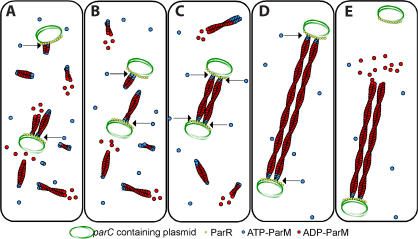
Type II par operons harness polymerization of the dynamically unstable actin-like protein ParM to segregate low-copy plasmids in rod-shaped bacteria. In this study, we use time-lapse fluorescence microscopy to follow plasmid dynamics and ParM assembly in Escherichia coli. Plasmids lacking a par operon undergo confined diffusion with a diffusion constant of 5 x 10(-5) microm(2)/s and a confinement radius of 0.28 microm. Single par-containing plasmids also move diffusively but with a larger diffusion constant (4 x 10(-4) microm(2)/s) and confinement radius (0.42 microm). ParM filaments are dynamically unstable in vivo and form spindles that link pairs of par-containing plasmids and drive them rapidly (3.1 microm/min) toward opposite poles of the cell. After reaching the poles, ParM filaments rapidly and completely depolymerize. After ParM disassembly, segregated plasmids resume diffusive motion, often encountering each other many times and undergoing multiple rounds of ParM-dependent segregation in a single cell cycle. We propose that in addition to driving segregation, the par operon enables plasmids to search space and find sister plasmids more effectively.
Figures
References
-
- Collins, J., and R.H. Pritchard. 1973. Relationship between chromosome replication and F'lac episome replication in Escherichia coli. J. Mol. Biol. 78:143–155. - PubMed
-
- Ebersbach, G., and K. Gerdes. 2005. Plasmid segregation mechanisms. Annu. Rev. Genet. 39:453–479. - PubMed
-
- Ebersbach, G., D.J. Sherratt, and K. Gerdes. 2005. Partition-associated incompatibility caused by random assortment of pure plasmid clusters. Mol. Microbiol. 56:1430–1440. - PubMed
-
- Garner, E.C., C.S. Campbell, and R.D. Mullins. 2004. Dynamic instability in a DNA-segregating prokaryotic actin homolog. Science. 306:1021–1025. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
