

Obligatory participation of macrophages in an angiopoietin 2-mediated cell death switch
- PMID: 18039971
- PMCID: PMC3675770
- DOI: 10.1242/dev.012187
Obligatory participation of macrophages in an angiopoietin 2-mediated cell death switch
Abstract
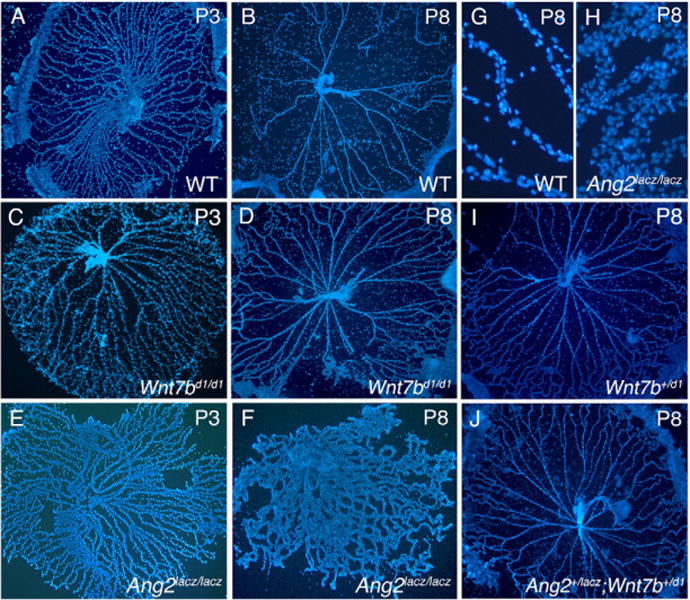
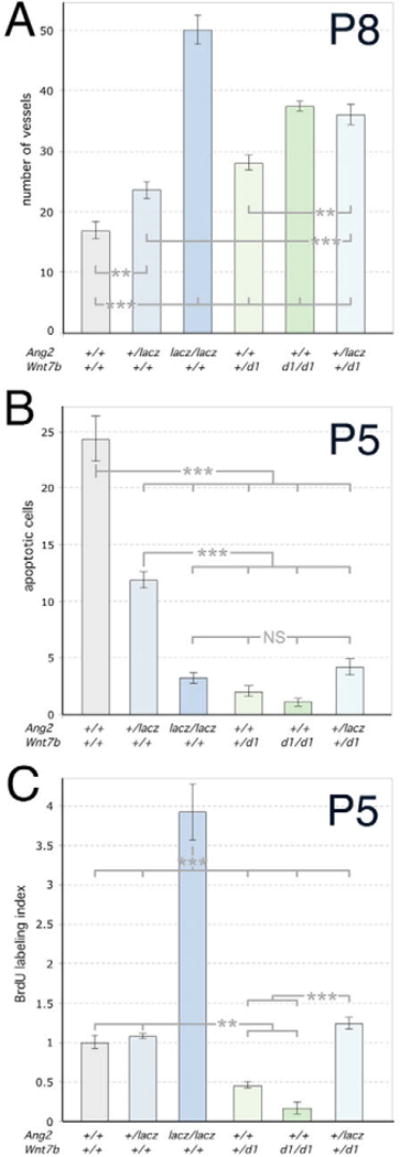
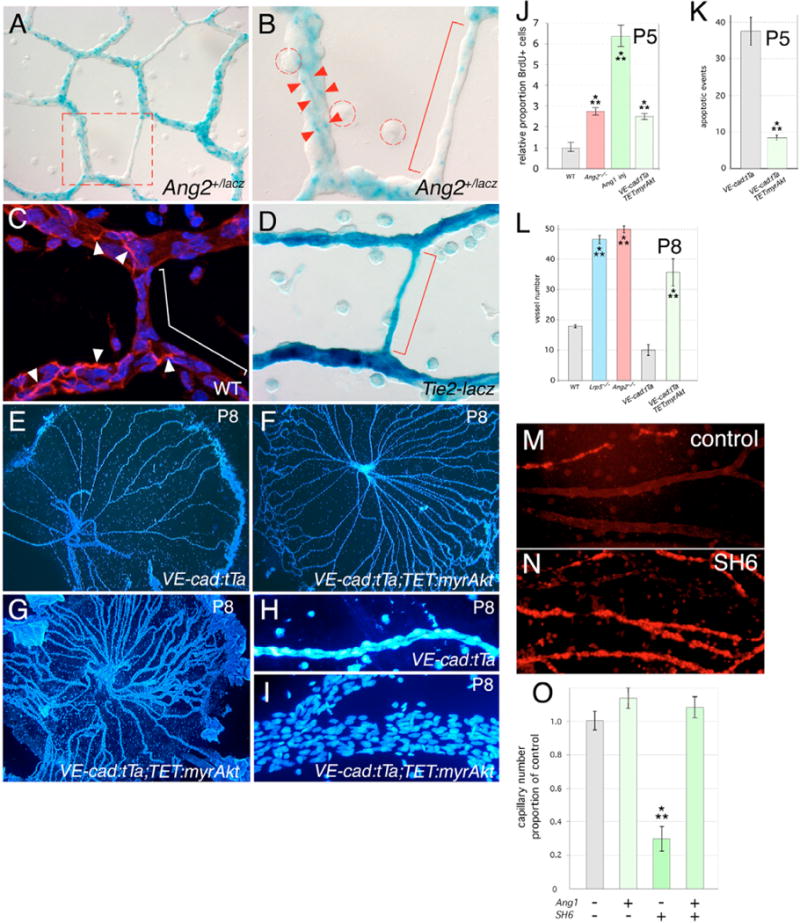
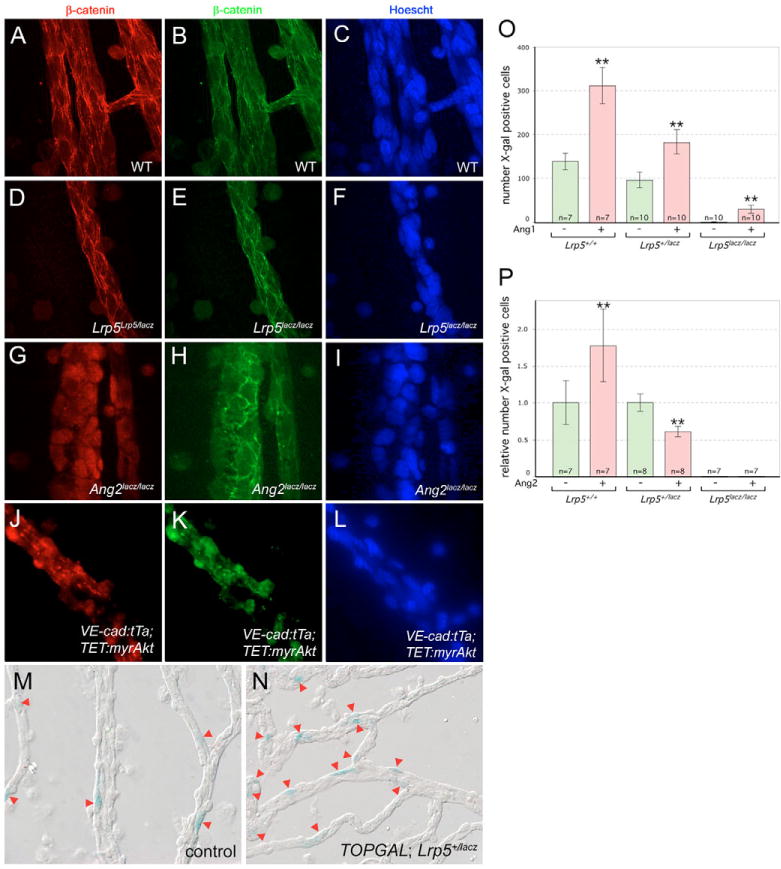
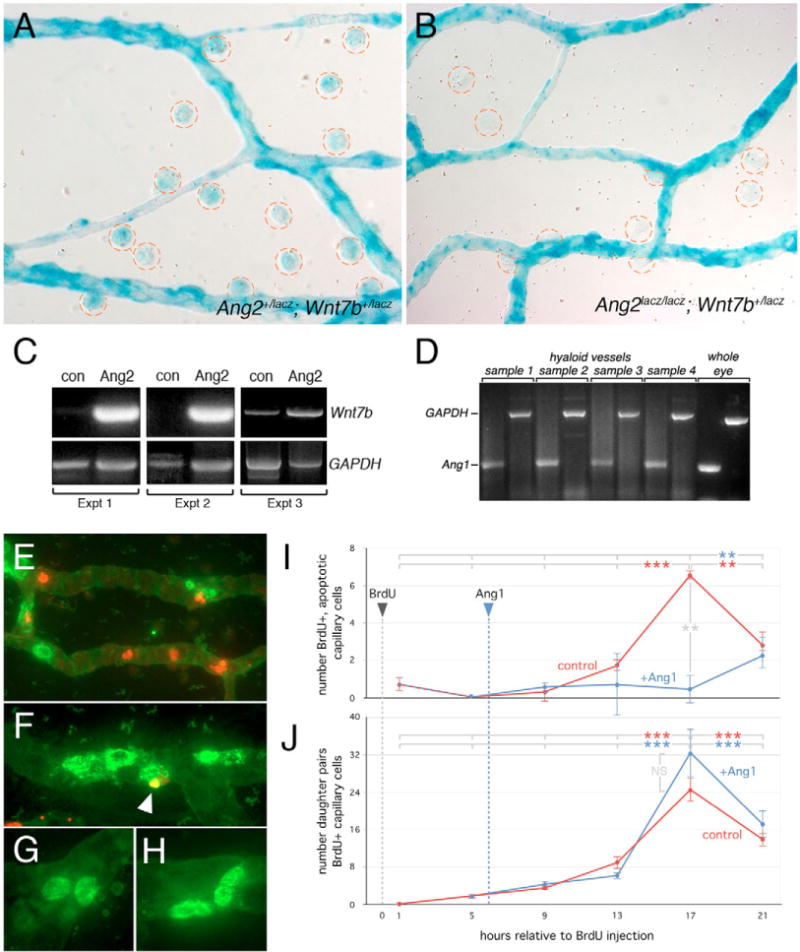
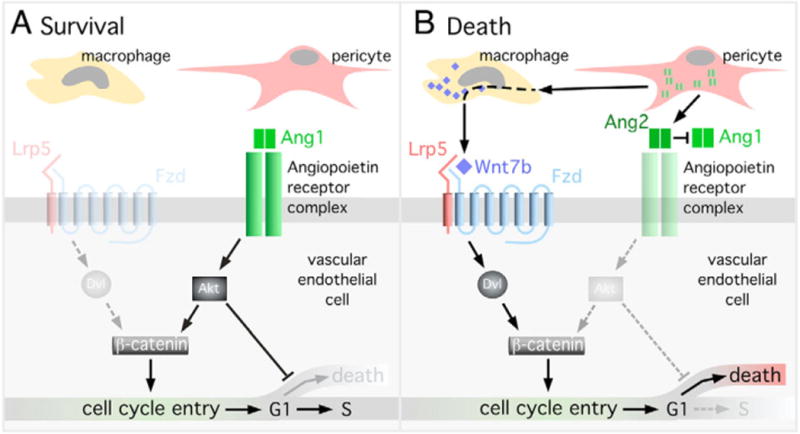
Macrophages have a critical function in the recognition and engulfment of dead cells. In some settings, macrophages also actively signal programmed cell death. Here we show that during developmentally scheduled vascular regression, resident macrophages are an obligatory participant in a signaling switch that favors death over survival. This switch occurs when the signaling ligand angiopoietin 2 has the dual effect of suppressing survival signaling in vascular endothelial cells (VECs) and stimulating Wnt ligand production by macrophages. In response to the Wnt ligand, VECs enter the cell cycle and in the absence of survival signals, die from G1 phase of the cell cycle. We propose that this mechanism represents an adaptation to ensure that the macrophage and its disposal capability are on hand when cell death occurs.
Figures

References
-
- Camenisch G, Pisabarro MT, Sherman D, Kowalski J, Nagel M, Hass P, Xie MH, Gurney A, Bodary S, Liang XH. ANGPTL3 stimulates endothelial cell adhesion and migration via integrin alpha vbeta 3 and induces blood vessel formation in vivo. J Biol Chem. 2002;277:17281–17290. - PubMed
-
- Carlson TR, Feng Y, Maisonpierre PC, Mrksich M, Morla AO. Direct cell adhesion to the angiopoietins mediated by integrins. J Biol Chem. 2001;276:26516–26525. - PubMed
-
- Castillo SS, Brognard J, Petukhov PA, Zhang C, Tsurutani J, Granville CA, Li M, Jung M, West KA, Gills JG. Preferential inhibition of Akt and killing of Akt-dependent cancer cells by rationally designed phosphatidylinositol ether lipid analogues. Cancer Res. 2004;64:2782–2792. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
