

Cryo-EM study of the spinach chloroplast ribosome reveals the structural and functional roles of plastid-specific ribosomal proteins
- PMID: 18042701
- PMCID: PMC2148287
- DOI: 10.1073/pnas.0709856104
Cryo-EM study of the spinach chloroplast ribosome reveals the structural and functional roles of plastid-specific ribosomal proteins
Abstract
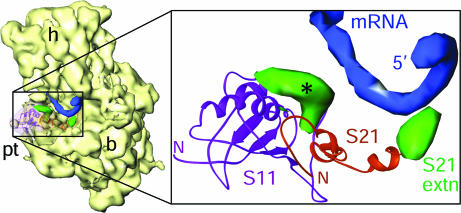
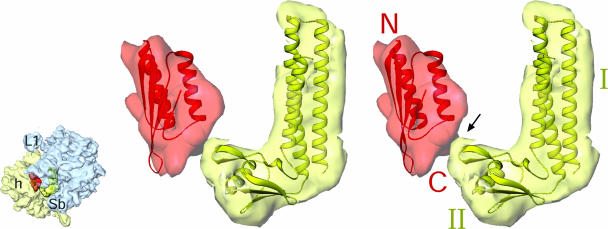
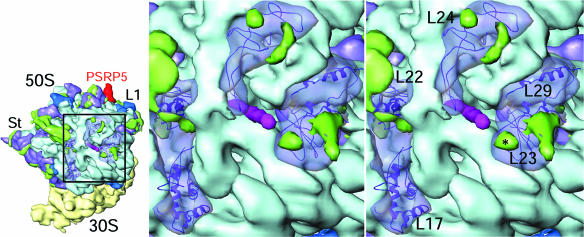
Protein synthesis in the chloroplast is carried out by chloroplast ribosomes (chloro-ribosome) and regulated in a light-dependent manner. Chloroplast or plastid ribosomal proteins (PRPs) generally are larger than their bacterial counterparts, and chloro-ribosomes contain additional plastid-specific ribosomal proteins (PSRPs); however, it is unclear to what extent these proteins play structural or regulatory roles during translation. We have obtained a three-dimensional cryo-EM map of the spinach 70S chloro-ribosome, revealing the overall structural organization to be similar to bacterial ribosomes. Fitting of the conserved portions of the x-ray crystallographic structure of the bacterial 70S ribosome into our cryo-EM map of the chloro-ribosome reveals the positions of PRP extensions and the locations of the PSRPs. Surprisingly, PSRP1 binds in the decoding region of the small (30S) ribosomal subunit, in a manner that would preclude the binding of messenger and transfer RNAs to the ribosome, suggesting that PSRP1 is a translation factor rather than a ribosomal protein. PSRP2 and PSRP3 appear to structurally compensate for missing segments of the 16S rRNA within the 30S subunit, whereas PSRP4 occupies a position buried within the head of the 30S subunit. One of the two PSRPs in the large (50S) ribosomal subunit lies near the tRNA exit site. Furthermore, we find a mass of density corresponding to chloro-ribosome recycling factor; domain II of this factor appears to interact with the flexible C-terminal domain of PSRP1. Our study provides evolutionary insights into the structural and functional roles that the PSRPs play during protein synthesis in chloroplasts.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- O'Brien TW. Gene. 2002;286:73–79. - PubMed
-
- Schmitz-Linneweber C, Maier RM, Alcaraz JP, Cottet A, Herrmann RG, Mache R. Plant Mol Biol. 2001;45:307–315. - PubMed
-
- Margulis L. Origin of Eukaryotic Cells. New Haven: Yale Univ Press; 1970.
-
- Yamaguchi K, Subramanian AR. J Biol Chem. 2000;275:28466–28482. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
