

Transforming growth factor: beta signaling is essential for limb regeneration in axolotls
- PMID: 18043735
- PMCID: PMC2082079
- DOI: 10.1371/journal.pone.0001227
Transforming growth factor: beta signaling is essential for limb regeneration in axolotls
Abstract
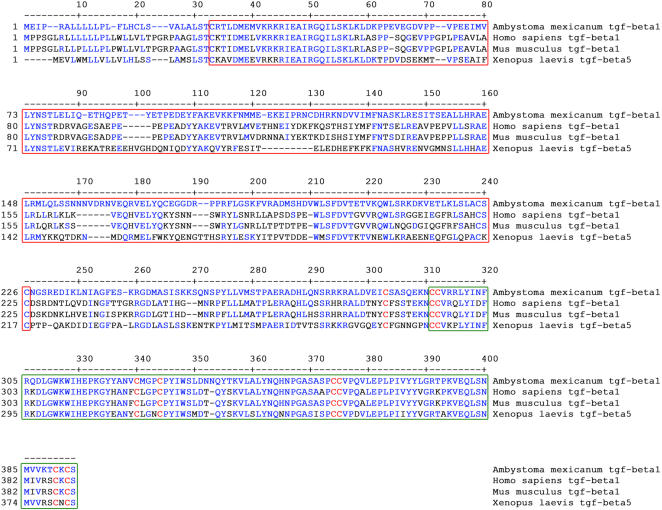
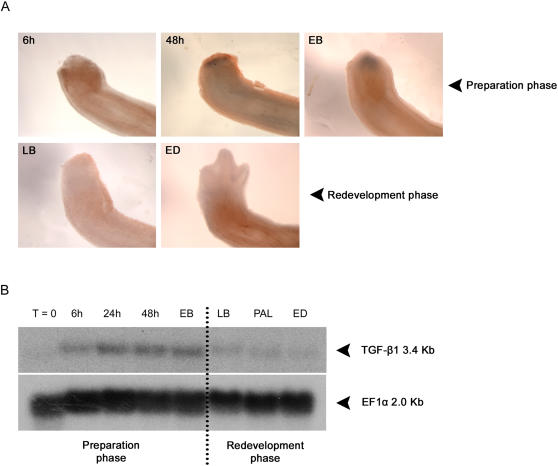
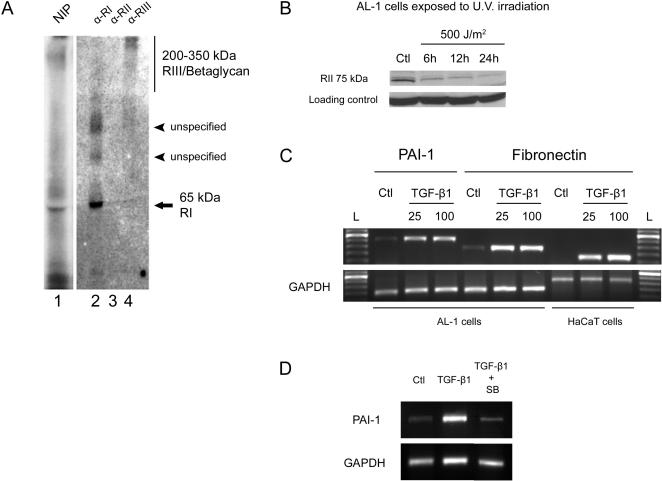
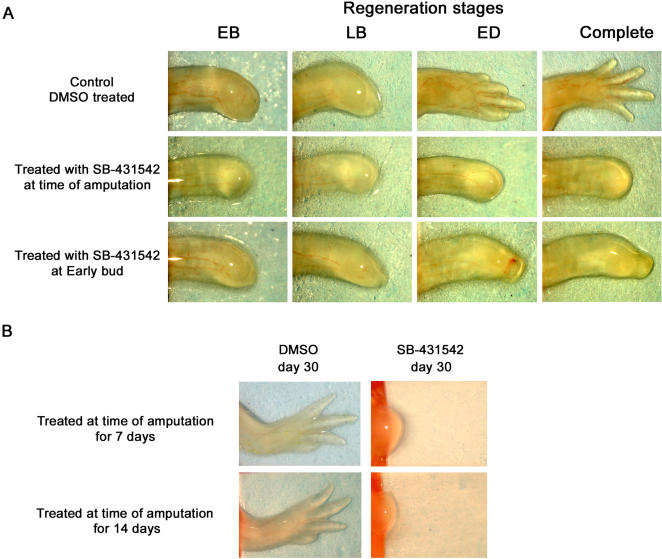
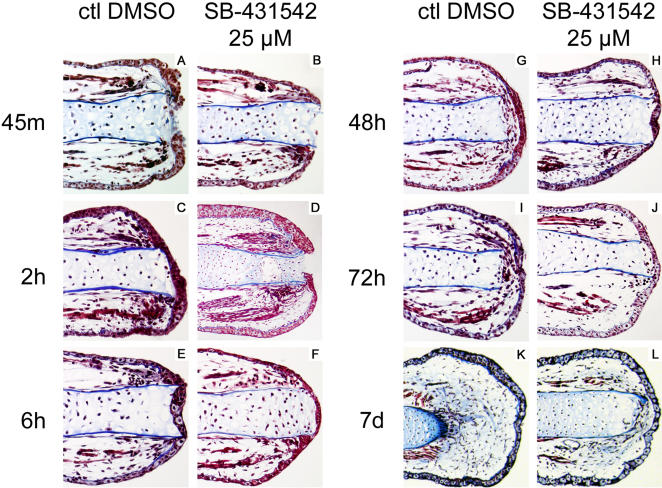
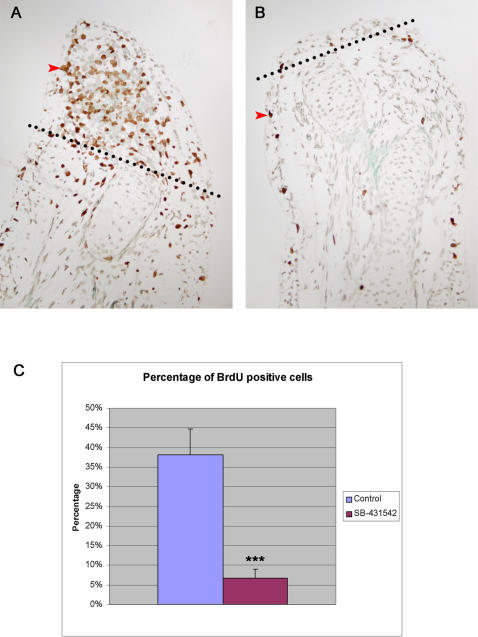
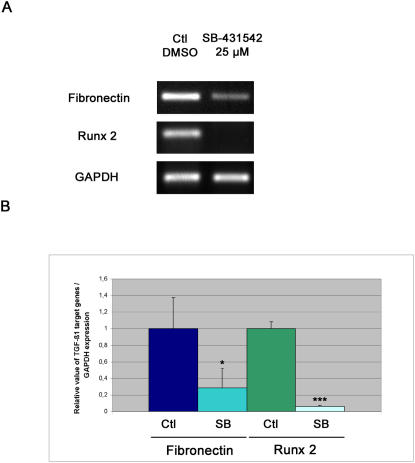
Axolotls (urodele amphibians) have the unique ability, among vertebrates, to perfectly regenerate many parts of their body including limbs, tail, jaw and spinal cord following injury or amputation. The axolotl limb is the most widely used structure as an experimental model to study tissue regeneration. The process is well characterized, requiring multiple cellular and molecular mechanisms. The preparation phase represents the first part of the regeneration process which includes wound healing, cellular migration, dedifferentiation and proliferation. The redevelopment phase represents the second part when dedifferentiated cells stop proliferating and redifferentiate to give rise to all missing structures. In the axolotl, when a limb is amputated, the missing or wounded part is regenerated perfectly without scar formation between the stump and the regenerated structure. Multiple authors have recently highlighted the similarities between the early phases of mammalian wound healing and urodele limb regeneration. In mammals, one very important family of growth factors implicated in the control of almost all aspects of wound healing is the transforming growth factor-beta family (TGF-beta). In the present study, the full length sequence of the axolotl TGF-beta1 cDNA was isolated. The spatio-temporal expression pattern of TGF-beta1 in regenerating limbs shows that this gene is up-regulated during the preparation phase of regeneration. Our results also demonstrate the presence of multiple components of the TGF-beta signaling machinery in axolotl cells. By using a specific pharmacological inhibitor of TGF-beta type I receptor, SB-431542, we show that TGF-beta signaling is required for axolotl limb regeneration. Treatment of regenerating limbs with SB-431542 reveals that cellular proliferation during limb regeneration as well as the expression of genes directly dependent on TGF-beta signaling are down-regulated. These data directly implicate TGF-beta signaling in the initiation and control of the regeneration process in axolotls.
Conflict of interest statement
Figures
Similar articles
-
Activation of Smad2 but not Smad3 is required to mediate TGF-β signaling during axolotl limb regeneration.Development. 2016 Oct 1;143(19):3481-3490. doi: 10.1242/dev.131466. Epub 2016 Aug 22. Development. 2016. PMID: 27549395
-
The axolotl limb: a model for bone development, regeneration and fracture healing.Bone. 2007 Jan;40(1):45-56. doi: 10.1016/j.bone.2006.07.005. Epub 2006 Aug 21. Bone. 2007. PMID: 16920050
-
Tgf-β superfamily and limb regeneration: Tgf-β to start and Bmp to end.Dev Dyn. 2022 Jun;251(6):973-987. doi: 10.1002/dvdy.379. Epub 2021 Jun 14. Dev Dyn. 2022. PMID: 34096672 Review.
-
Skin wound healing in axolotls: a scarless process.J Exp Zool B Mol Dev Evol. 2010 Dec 15;314(8):684-97. doi: 10.1002/jez.b.21371. J Exp Zool B Mol Dev Evol. 2010. PMID: 20718005
-
Identification of immune and non-immune cells in regenerating axolotl limbs by single-cell sequencing.Exp Cell Res. 2020 Sep 15;394(2):112149. doi: 10.1016/j.yexcr.2020.112149. Epub 2020 Jun 18. Exp Cell Res. 2020. PMID: 32562784 Free PMC article. Review.
Cited by
-
The optimization of PLGA knitted mesh reinforced-collagen/chitosan scaffold for the healing of full-thickness skin defects.J Biomed Mater Res B Appl Biomater. 2023 Apr;111(4):763-774. doi: 10.1002/jbm.b.35187. Epub 2022 Nov 11. J Biomed Mater Res B Appl Biomater. 2023. PMID: 36367718 Free PMC article.
-
Evaluation of the anesthetic effects of MS222 in the adult Mexican axolotl (Ambystoma mexicanum).Vet Med (Auckl). 2016 Jan 5;7:1-7. doi: 10.2147/VMRR.S96761. eCollection 2016. Vet Med (Auckl). 2016. PMID: 30050832 Free PMC article.
-
The Regulation of Growth in Developing, Homeostatic, and Regenerating Tetrapod Limbs: A Minireview.Front Cell Dev Biol. 2022 Jan 3;9:768505. doi: 10.3389/fcell.2021.768505. eCollection 2021. Front Cell Dev Biol. 2022. PMID: 35047496 Free PMC article. Review.
-
Digit Tip Injuries: Current Treatment and Future Regenerative Paradigms.Stem Cells Int. 2019 Jan 21;2019:9619080. doi: 10.1155/2019/9619080. eCollection 2019. Stem Cells Int. 2019. PMID: 30805012 Free PMC article. Review.
-
Mapping hematopoiesis in a fully regenerative vertebrate: the axolotl.Blood. 2014 Aug 21;124(8):1232-41. doi: 10.1182/blood-2013-09-526970. Epub 2014 May 6. Blood. 2014. PMID: 24802774 Free PMC article.
References
-
- Wallace H. New York: Wiley; 1981. Vertebrate limb regeneration. Chichester Eng.; p. xii, 276.
-
- Tsonis PA. New York, NY, USA: Cambridge University Press; 1996. Limb regeneration. Cambridge [England]; p. xii, 241.
-
- Goss RJ. Prospects of regeneration in man. Clin Orthop Relat Res. 1980:270–282. - PubMed
-
- Martin P. Wound healing–aiming for perfect skin regeneration. Science. 1997;276:75–81. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
