

Resolving the fast kinetics of cooperative binding: Ca2+ buffering by calretinin
- PMID: 18044987
- PMCID: PMC2229850
- DOI: 10.1371/journal.pbio.0050311
Resolving the fast kinetics of cooperative binding: Ca2+ buffering by calretinin
Abstract
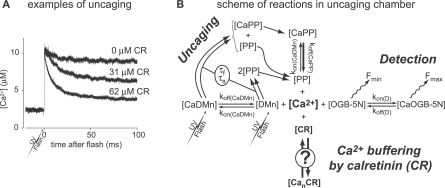
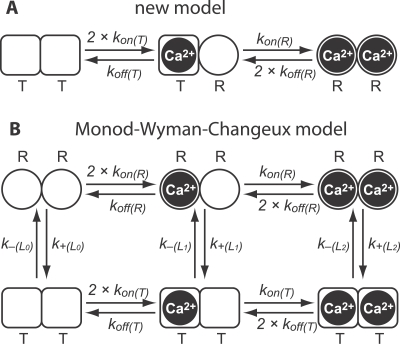
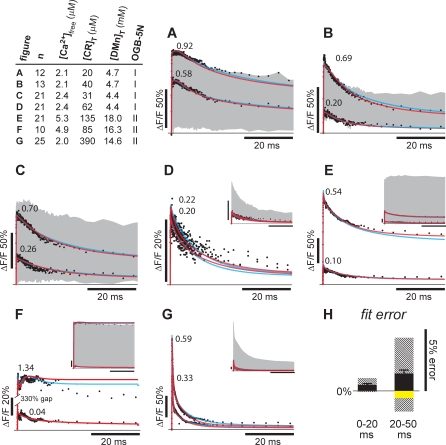
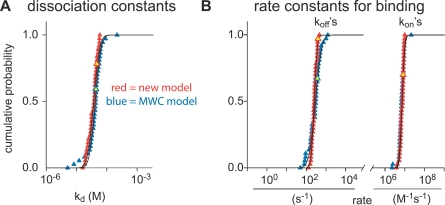
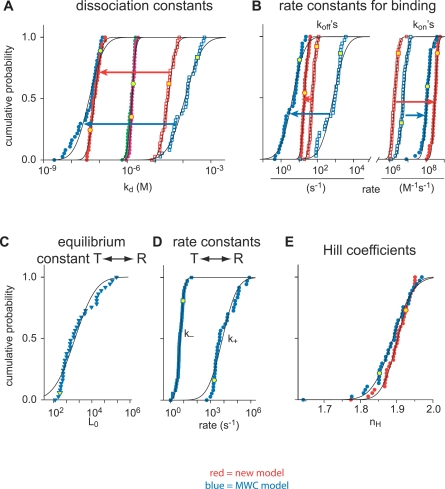
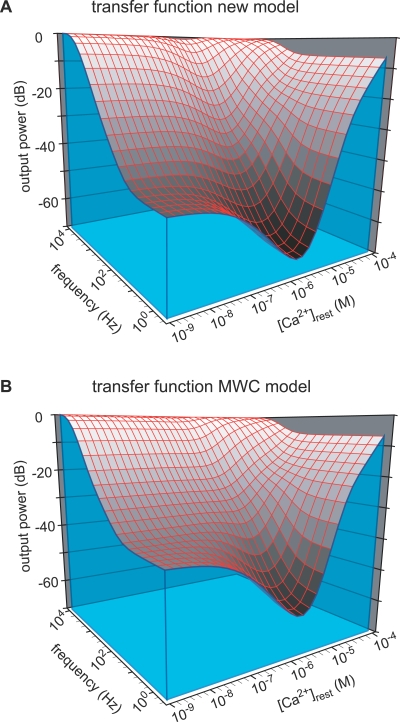
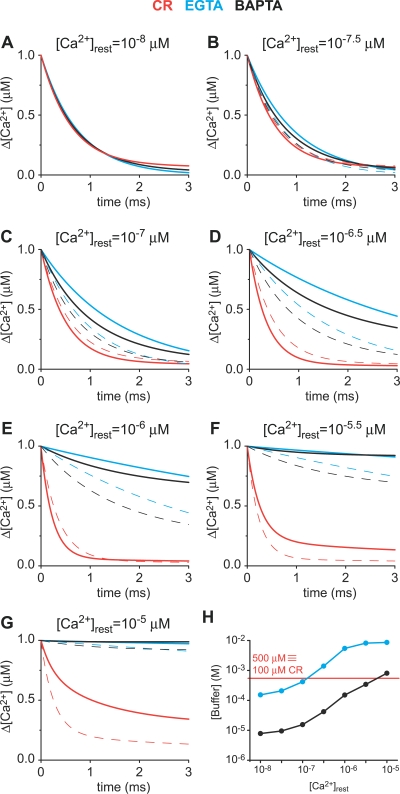
Cooperativity is one of the most important properties of molecular interactions in biological systems. It is the ability to influence ligand binding at one site of a macromolecule by previous ligand binding at another site of the same molecule. As a consequence, the affinity of the macromolecule for the ligand is either decreased (negative cooperativity) or increased (positive cooperativity). Over the last 100 years, O2 binding to hemoglobin has served as the paradigm for cooperative ligand binding and allosteric modulation, and four practical models were developed to quantitatively describe the mechanism: the Hill, the Adair-Klotz, the Monod-Wyman-Changeux, and the Koshland-Némethy-Filmer models. The predictions of these models apply under static conditions when the binding reactions are at equilibrium. However, in a physiological setting, e.g., inside a cell, the timing and dynamics of the binding events are essential. Hence, it is necessary to determine the dynamic properties of cooperative binding to fully understand the physiological implications of cooperativity. To date, the Monod-Wyman-Changeux model was applied to determine the kinetics of cooperative binding to biologically active molecules. In this model, cooperativity is established by postulating two allosteric isoforms with different binding properties. However, these studies were limited to special cases, where transition rates between allosteric isoforms are much slower than the binding rates or where binding and unbinding rates could be measured independently. For all other cases, the complex mathematical description precludes straightforward interpretations. Here, we report on calculating for the first time the fast dynamics of a cooperative binding process, the binding of Ca2+ to calretinin. Calretinin is a Ca2+-binding protein with four cooperative binding sites and one independent binding site. The Ca2+ binding to calretinin was assessed by measuring the decay of free Ca2+ using a fast fluorescent Ca2+ indicator following rapid (<50-mus rise time) Ca2+ concentration jumps induced by uncaging Ca2+ from DM-nitrophen. To unravel the kinetics of cooperative binding, we devised several approaches based on known cooperative binding models, resulting in a novel and relatively simple model. This model revealed unexpected and highly specific nonlinear properties of cellular Ca2+ regulation by calretinin. The association rate of Ca2+ with calretinin speeds up as the free Ca2+ concentration increases from cytoplasmic resting conditions ( approximately 100 nM) to approximately 1 muM. As a consequence, the Ca2+ buffering speed of calretinin highly depends on the prevailing Ca2+ concentration prior to a perturbation. In addition to providing a novel mode of action of cellular Ca2+ buffering, our model extends the analysis of cooperativity beyond the static steady-state condition, providing a powerful tool for the investigation of the dynamics and functional significance of cooperative binding in general.
Conflict of interest statement
Figures
References
-
- Zhang M, Yuan T. Molecular mechanisms of calmodulin's functional versatility. Biochem Cell Biol. 1998;76:313–323. - PubMed
-
- Schwaller B, Durussel I, Jermann D, Herrmann B, Cox JA. Comparison of the Ca2+-binding properties of human recombinant calretinin-22k and calretinin. J Biol Chem. 1997;272:29663–29671. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
