

EGF and amphiregulin differentially regulate Cbl recruitment to endosomes and EGF receptor fate
- PMID: 18045238
- PMCID: PMC3507514
- DOI: 10.1042/BJ20071505
EGF and amphiregulin differentially regulate Cbl recruitment to endosomes and EGF receptor fate
Abstract
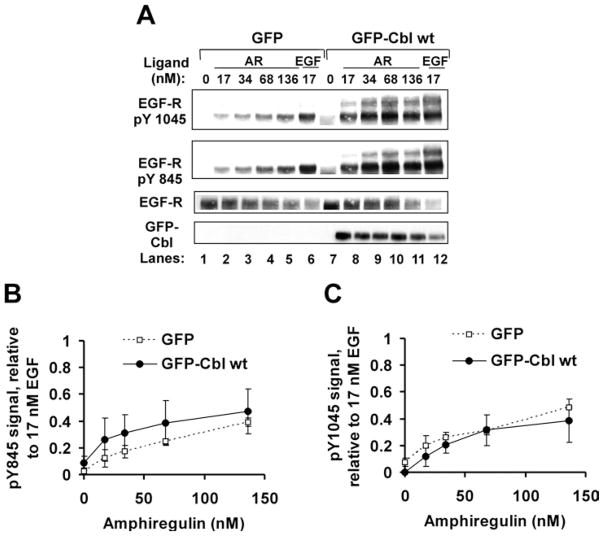
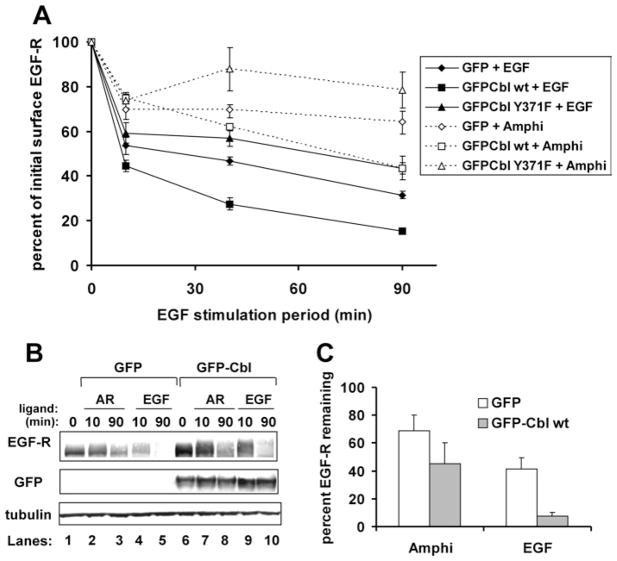
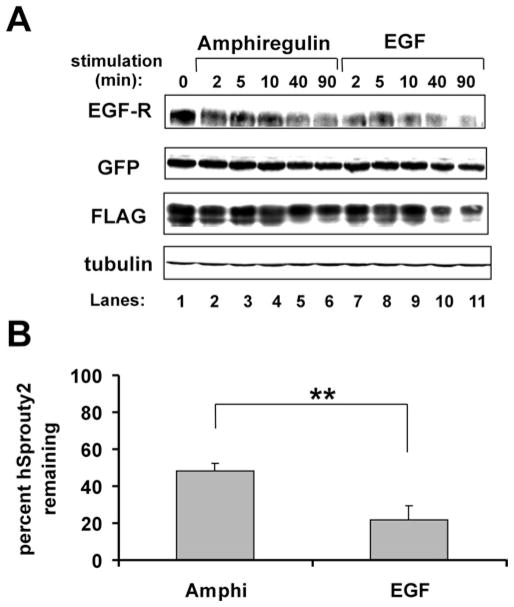
EGF-R [EGF (epidermal growth factor) receptor] ligands can promote or inhibit cell growth. The biological outcome of receptor activation is dictated, at least in part, by ligand-specified patterns of endocytic trafficking. EGF-R trafficking downstream of the ligands EGF and TGF-alpha (transforming growth factor-alpha) has been investigated extensively. However, less is known about EGF-R fates induced by the ligands BTC (betacellulin) and AR (amphiregulin). We undertook comparative analyses to identify ligand-specific molecular events that regulate EGF-R trafficking and degradation. EGF (17 nM) and BTC (8.5 nM) induced significant EGF-R degradation, with or without ectopic expression of the ubiquitin ligase Cbl. Human recombinant AR (17 nM) failed to affect receptor degradation in either case. Notably, levels of ligand-induced EGF-R ubiquitination did not correlate strictly with receptor degradation. Dose-response experiments revealed that AR at a saturating concentration was a partial agonist at the EGF-R, with approx. 40% efficacy (relative to EGF) at inducing receptor tyrosine phosphorylation, ubiquitination and association with Cbl. EGF-R down-regulation and degradation also were compromised upon cell stimulation with AR (136 nM). These outcomes correlated with decreased degradation of the Cbl substrate and internalization inhibitor hSprouty2. Downstream of the hSprouty2 checkpoint in AR-stimulated cells, Cbl-free EGF-R was incorporated into endosomes from which Cbl-EGF-R complexes were excluded. Our results suggest that the AR-specific EGF-R fate results from decreased hSprouty2 degradation and reduced Cbl recruitment to underphosphorylated EGF-R, two effects that impair EGF-R trafficking to lysosomes.
Figures



References
-
- Luetteke NC, Qiu TH, Fenton SE, Troyer KL, Riedel RF, Chang A, Lee DC. Targeted inactivation of the EGF and amphiregulin genes reveals distinct roles for EGF receptor ligands in mouse mammary gland development. Development. 1999;126:2739–2750. - PubMed
-
- Xian CJ, Li L, Deng YS, Zhao SP, Zhou XF. Lack of effects of transforming growth factor-α gene knockout on peripheral nerve regeneration may result from compensatory mechanisms. Exp Neurol. 2001;172:182–188. - PubMed
-
- Abbott BD, Lin TM, Rasmussen NT, Albrecht RM, Schmid JE, Peterson RE. Lack of expression of EGF and TGF-α in the fetal mouse alters formation of prostatic epithelial buds and influences the response to TCDD. Toxicol Sci. 2003;76:427–436. - PubMed
-
- Troyer KL, Luetteke NC, Saxon ML, Qiu TH, Xian CJ, Lee DC. Growth retardation, duodenal lesions, and aberrant ileum architecture in triple null mice lacking EGF, amphiregulin, and TGF-α. Gastroenterology. 2001;121:68–78. - PubMed
-
- Luetteke NC, Qiu TH, Peiffer RL, Oliver P, Smithies O, Lee DC. TGFα deficiency results in hair follicle and eye abnormalities in targeted and waved-1 mice. Cell. 1993;73:263–278. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous