

Myosin transducer mutations differentially affect motor function, myofibril structure, and the performance of skeletal and cardiac muscles
- PMID: 18045988
- PMCID: PMC2230588
- DOI: 10.1091/mbc.e07-09-0890
Myosin transducer mutations differentially affect motor function, myofibril structure, and the performance of skeletal and cardiac muscles
Abstract
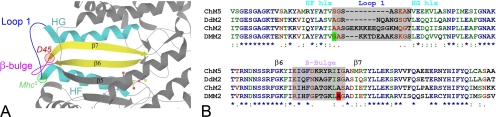
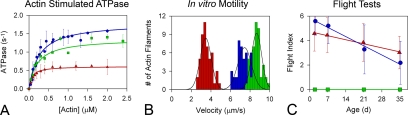
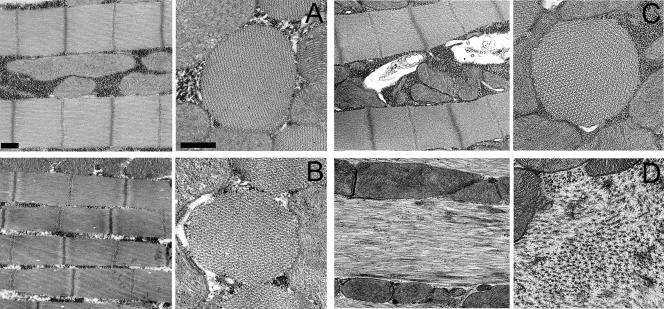
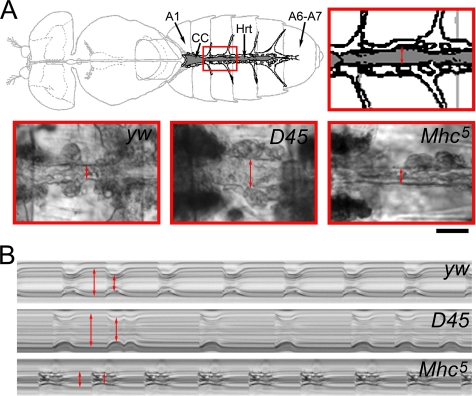
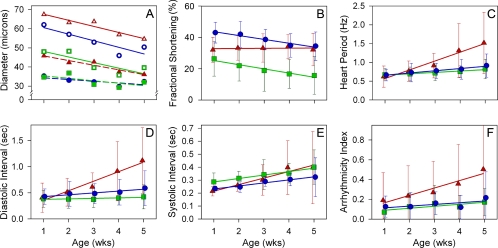
Striated muscle myosin is a multidomain ATP-dependent molecular motor. Alterations to various domains affect the chemomechanical properties of the motor, and they are associated with skeletal and cardiac myopathies. The myosin transducer domain is located near the nucleotide-binding site. Here, we helped define the role of the transducer by using an integrative approach to study how Drosophila melanogaster transducer mutations D45 and Mhc(5) affect myosin function and skeletal and cardiac muscle structure and performance. We found D45 (A261T) myosin has depressed ATPase activity and in vitro actin motility, whereas Mhc(5) (G200D) myosin has these properties enhanced. Depressed D45 myosin activity protects against age-associated dysfunction in metabolically demanding skeletal muscles. In contrast, enhanced Mhc(5) myosin function allows normal skeletal myofibril assembly, but it induces degradation of the myofibrillar apparatus, probably as a result of contractile disinhibition. Analysis of beating hearts demonstrates depressed motor function evokes a dilatory response, similar to that seen with vertebrate dilated cardiomyopathy myosin mutations, and it disrupts contractile rhythmicity. Enhanced myosin performance generates a phenotype apparently analogous to that of human restrictive cardiomyopathy, possibly indicating myosin-based origins for the disease. The D45 and Mhc(5) mutations illustrate the transducer's role in influencing the chemomechanical properties of myosin and produce unique pathologies in distinct muscles. Our data suggest Drosophila is a valuable system for identifying and modeling mutations analogous to those associated with specific human muscle disorders.
Figures
References
-
- Ahmad F., Seidman J. G., Seidman C. E. The genetic basis for cardiac remodeling. Annu. Rev. Genomics Hum. Genet. 2005;6:185–216. - PubMed
-
- Baker G. T., 3rd Insect flight muscle: maturation and senescence. Gerontology. 1976;22:334–361. - PubMed
-
- Bernstein S. I., Mogami K., Donady J. J., Emerson C. P., Jr Drosophila muscle myosin heavy chain encoded by a single gene in a cluster of muscle mutations. Nature. 1983;302:393–397. - PubMed
-
- Brown J. H., Cohen C. Regulation of muscle contraction by tropomyosin and troponin: how structure illuminates function. Adv. Protein Chem. 2005;71:121–159. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
