

RNA-mediated epigenetic programming of a genome-rearrangement pathway
- PMID: 18046331
- PMCID: PMC2647009
- DOI: 10.1038/nature06452
RNA-mediated epigenetic programming of a genome-rearrangement pathway
Abstract
Genome-wide DNA rearrangements occur in many eukaryotes but are most exaggerated in ciliates, making them ideal model systems for epigenetic phenomena. During development of the somatic macronucleus, Oxytricha trifallax destroys 95% of its germ line, severely fragmenting its chromosomes, and then unscrambles hundreds of thousands of remaining fragments by permutation or inversion. Here we demonstrate that DNA or RNA templates can orchestrate these genome rearrangements in Oxytricha, supporting an epigenetic model for sequence-dependent comparison between germline and somatic genomes. A complete RNA cache of the maternal somatic genome may be available at a specific stage during development to provide a template for correct and precise DNA rearrangement. We show the existence of maternal RNA templates that could guide DNA assembly, and that disruption of specific RNA molecules disables rearrangement of the corresponding gene. Injection of artificial templates reprogrammes the DNA rearrangement pathway, suggesting that RNA molecules guide genome rearrangement.
Figures
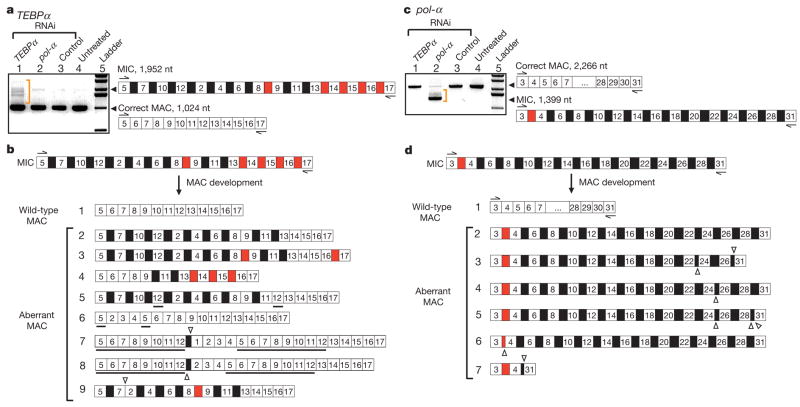
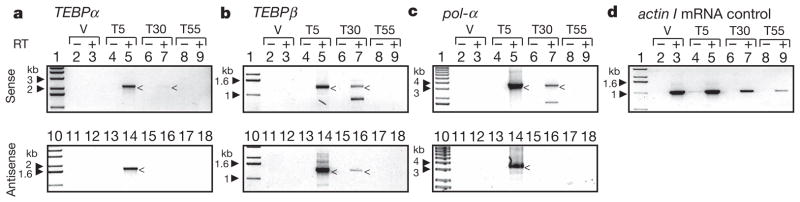

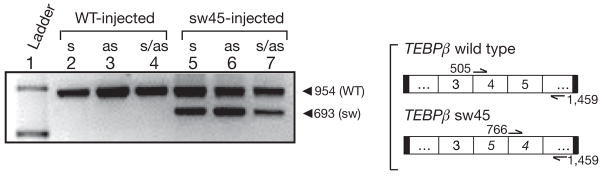
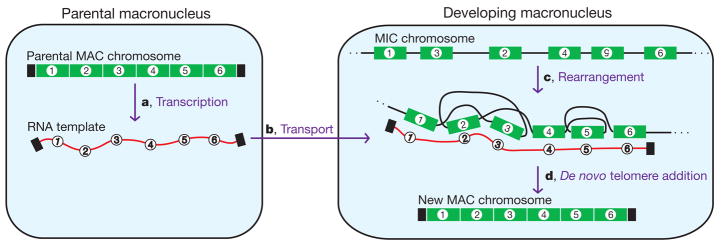
Comment in
-
Molecular biology. RNA rules.Nature. 2008 Jan 10;451(7175):131-2. doi: 10.1038/451131a. Nature. 2008. PMID: 18185572 No abstract available.
References
-
- King ML, Messitt TJ, Mowry KL. Putting RNAs in the right place at the right time: RNA localization in the frog oocyte. Biol Cell. 2005;97:19–33. - PubMed
-
- Tadros W, Lipshitz HD. Setting the stage for development: mRNA translation and stability during oocyte maturation and egg activation in Drosophila. Dev Dyn. 2005;232:593–608. - PubMed
-
- Rassoulzadegan M, et al. RNA-mediated non-mendelian inheritance of an epigenetic change in the mouse. Nature. 2006;441:469–474. - PubMed
-
- Herbert A, Rich A. RNA processing and the evolution of eukaryotes. Nature Genet. 1999;21:265–269. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
