

Calcineurin sets the bandwidth for discrimination of signals during thymocyte development
- PMID: 18046413
- PMCID: PMC5336393
- DOI: 10.1038/nature06305
Calcineurin sets the bandwidth for discrimination of signals during thymocyte development
Abstract
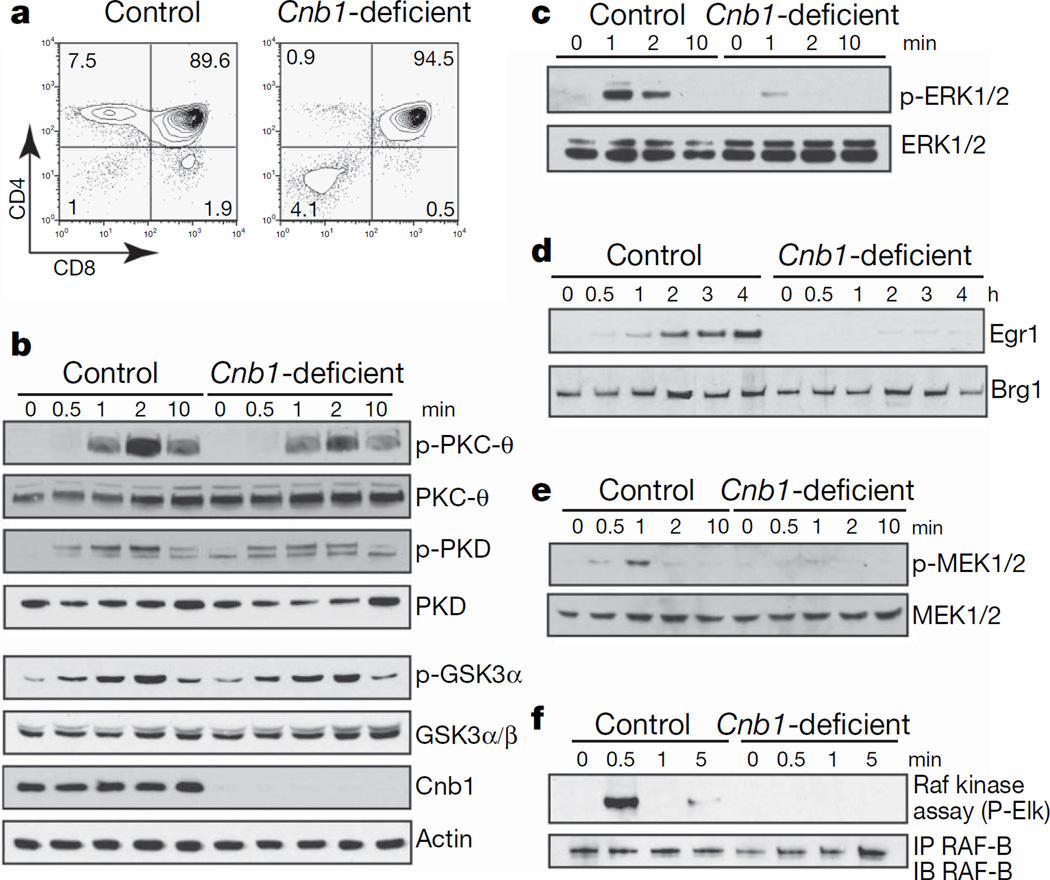
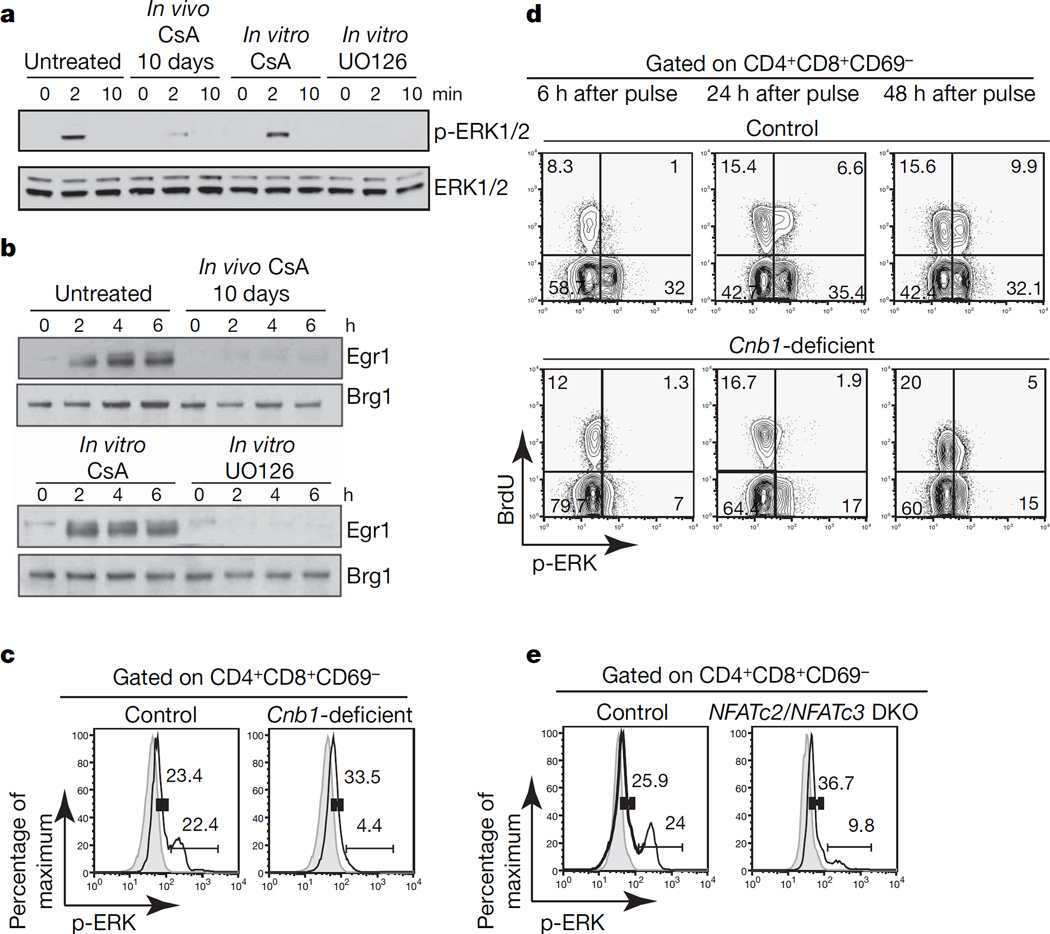
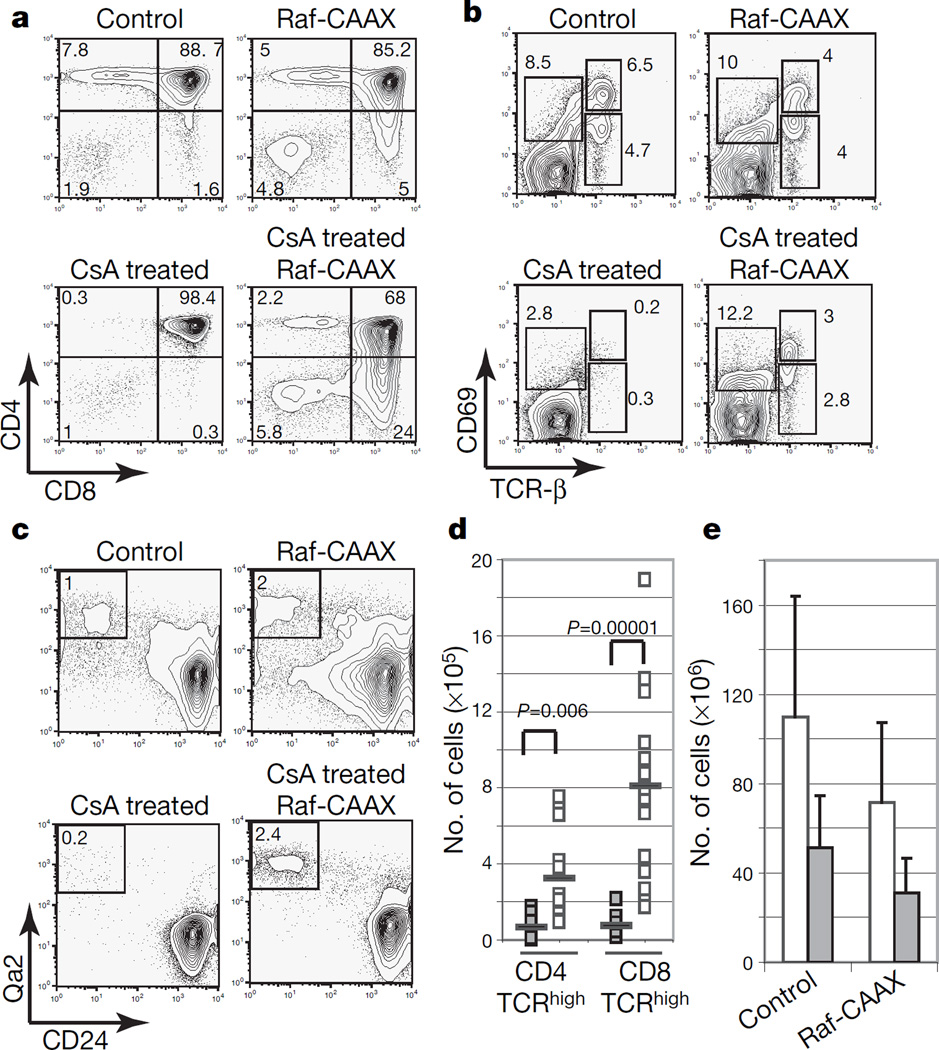
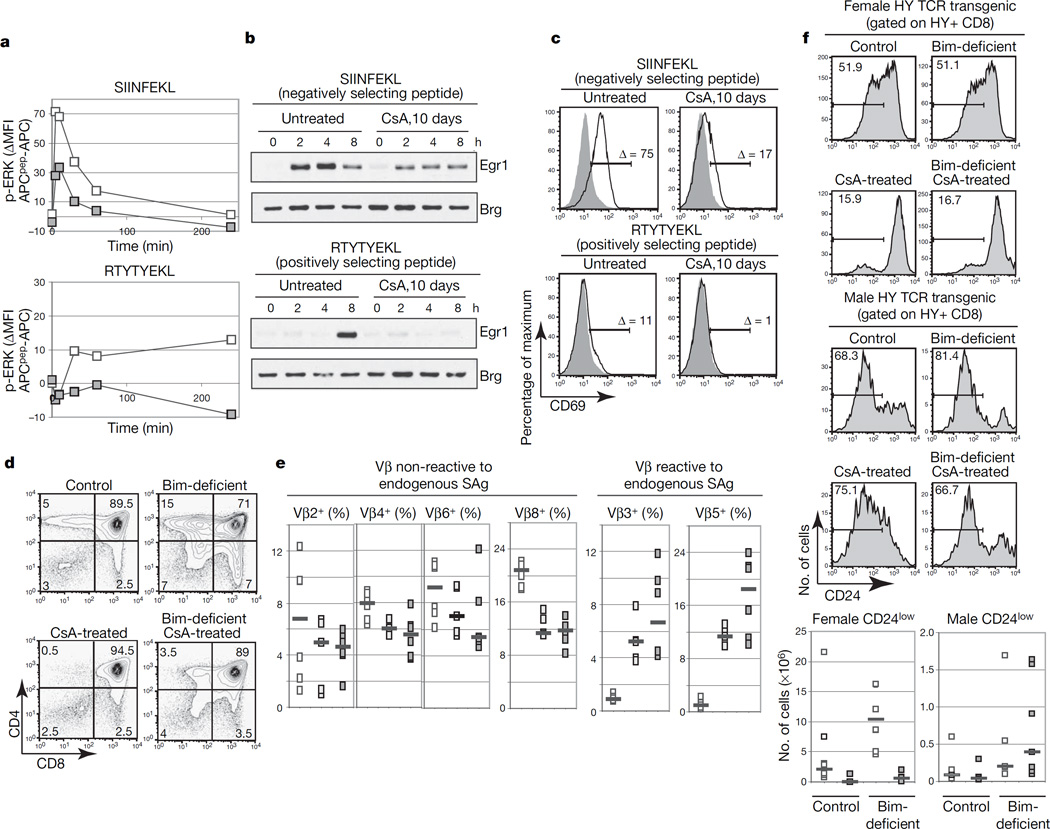
At critical times in development, cells are able to convert graded signals into discrete developmental outcomes; however, the mechanisms involved are poorly understood. During thymocyte development, cell fate is determined by signals originating from the alphabeta T-cell receptor. Low-affinity/avidity interactions between the T-cell receptor and peptide-MHC complexes direct differentiation to the single-positive stage (positive selection), whereas high-affinity/avidity interactions induce death by apoptosis (negative selection). Here we show that mice deficient in both calcineurin and nuclear factor of activated T cells (NFAT)c2/c3 lack a population of preselection thymocytes with enhanced ability to activate the mitogen-activated protein kinase (Raf-MEK-ERK) pathway, and fail to undergo positive selection. This defect can be partially rescued with constitutively active Raf, indicating that calcineurin controls MAPK signalling. Analysis of mice deficient in both Bim (which is required for negative selection) and calcineurin revealed that calcineurin-induced ERK (extracellular signal-regulated kinase) sensitization is required for differentiation in response to 'weak' positive selecting signals but not in response to 'strong' negative selecting signals (which normally induce apoptosis). These results indicate that early calcineurin/NFAT signalling produces a developmental period of ERK hypersensitivity, allowing very weak signals to induce positive selection. This mechanism might be generally useful in the discrimination of graded signals that induce different cell fates.
Figures
References
-
- Palmer E. Negative selection–clearing out the bad apples from the T-cell repertoire. Nature Rev. Immunol. 2003;3:383–391. - PubMed
-
- Starr TK, Jameson SC, Hogquist KA. Positive and negative selection of T cells. Annu. Rev. Immunol. 2003;21:139–176. - PubMed
-
- Neilson JR, Winslow MM, Hur EM, Crabtree GR. Calcineurin B1 is essential for positive but not negative selection during thymocyte development. Immunity. 2004;20:255–266. - PubMed
-
- Cante-Barrett K, Winslow MM, Crabtree GR. Selective role of NFATc3 in positive selection of thymocytes. J. Immunol. 2007;179:103–110. - PubMed
-
- Alberola-Ila J, Forbush KA, Seger R, Krebs EG, Perlmutter RM. Selective requirement for MAP kinase activation in thymocyte differentiation. Nature. 1995;373:620–623. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
