

Crystal structure of human XLF/Cernunnos reveals unexpected differences from XRCC4 with implications for NHEJ
- PMID: 18046455
- PMCID: PMC2104711
- DOI: 10.1038/sj.emboj.7601942
Crystal structure of human XLF/Cernunnos reveals unexpected differences from XRCC4 with implications for NHEJ
Abstract
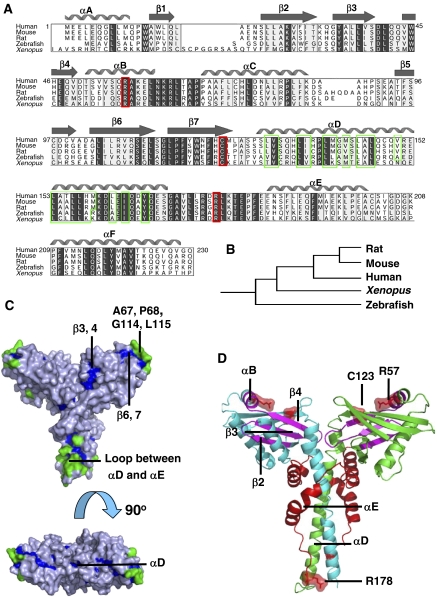
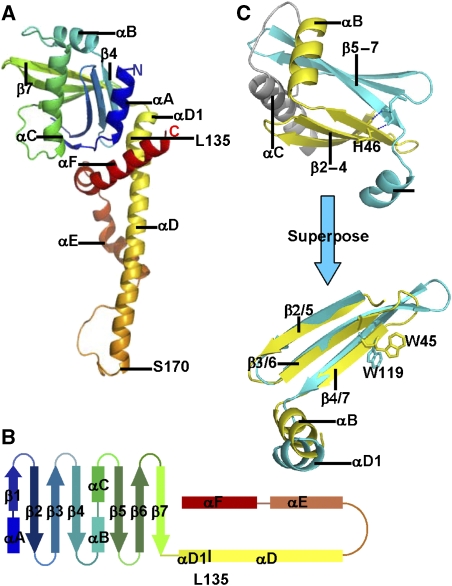
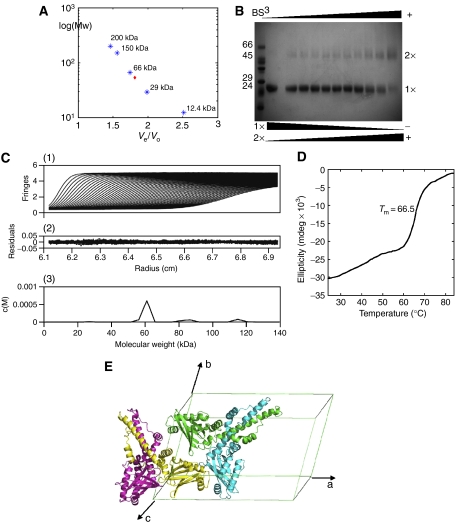
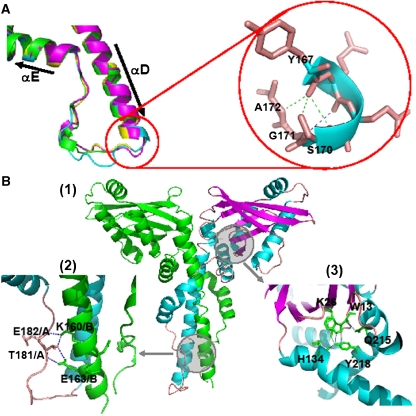
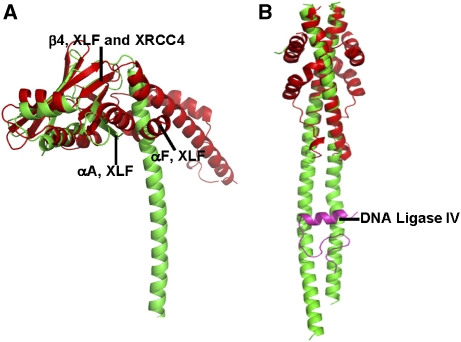
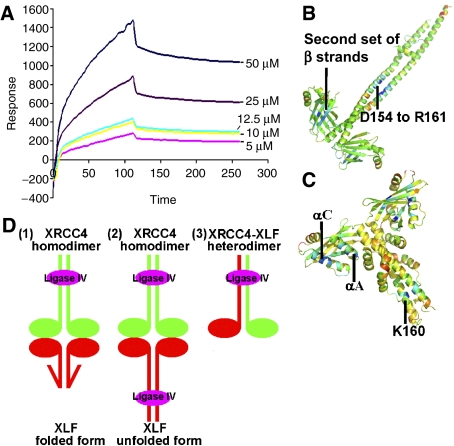
The recently characterised 299-residue human XLF/Cernunnos protein plays a crucial role in DNA repair by non-homologous end joining (NHEJ) and interacts with the XRCC4-DNA Ligase IV complex. Here, we report the crystal structure of the XLF (1-233) homodimer at 2.3 A resolution, confirming the predicted structural similarity to XRCC4. The XLF coiled-coil, however, is shorter than that of XRCC4 and undergoes an unexpected reverse in direction giving rise to a short distorted four helical bundle and a C-terminal helical structure wedged between the coiled-coil and head domain. The existence of a dimer as the major species is confirmed by size-exclusion chromatography, analytical ultracentrifugation, small-angle X-ray scattering and other biophysical methods. We show that the XLF structure is not easily compatible with a proposed XRCC4:XLF heterodimer. However, we demonstrate interactions between dimers of XLF and XRCC4 by surface plasmon resonance and analyse these in terms of surface properties, amino-acid conservation and mutations in immunodeficient patients. Our data are most consistent with head-to-head interactions in a 2:2:1 XRCC4:XLF:Ligase IV complex.
Figures
References
-
- Ahnesorg P, Smith P, Jackson SP (2006) XLF interacts with the XRCC4–DNA ligase IV complex to promote DNA nonhomologous end-joining. Cell 124: 301–313 - PubMed
-
- Buck D, Malivert L, de Chasseval R, Barraud A, Fondaneche MC, Sanal O, Plebani A, Stephan JL, Hufnagel M, le Deist F, Fischer A, Durandy A, de Villartay JP, Revy P (2006) Cernunnos, a novel nonhomologous end-joining factor, is mutated in human immunodeficiency with microcephaly. Cell 124: 287–299 - PubMed
-
- Callebaut I, Malivert L, Fischer A, Mornon JP, Revy P, de Villartay JP (2006) Cernunnos interacts with the XRCC4 x DNA–ligase IV complex and is homologous to the yeast nonhomologous end-joining factor Nej1. J Biol Chem 281: 13857–13860 - PubMed
-
- Critchlow SE, Bowater RP, Jackson SP (1997) Mammalian DNA double-strand break repair protein XRCC4 interacts with DNA ligase IV. Curr Biol 7: 588–598 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
