

Structure-guided identification of a new catalytic motif of oligosaccharyltransferase
- PMID: 18046457
- PMCID: PMC2206122
- DOI: 10.1038/sj.emboj.7601940
Structure-guided identification of a new catalytic motif of oligosaccharyltransferase
Abstract
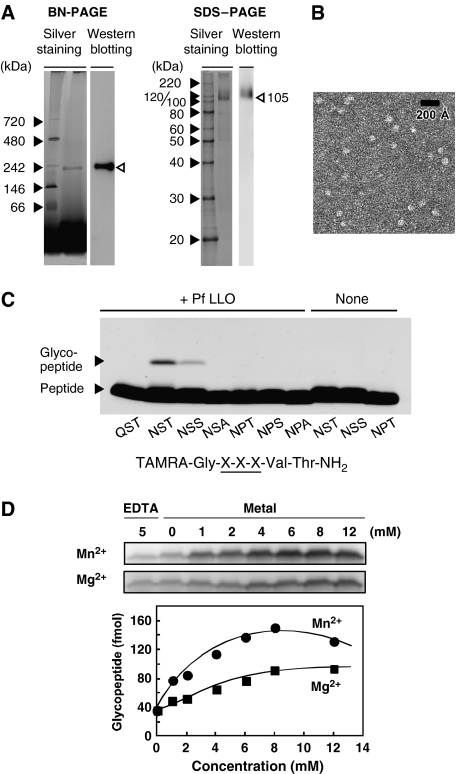
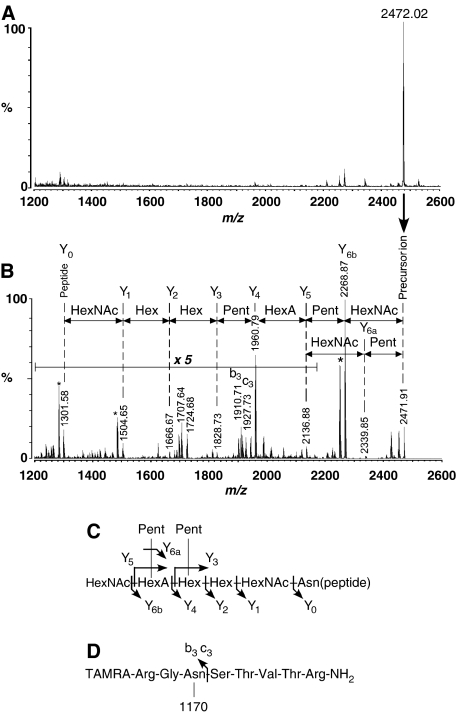
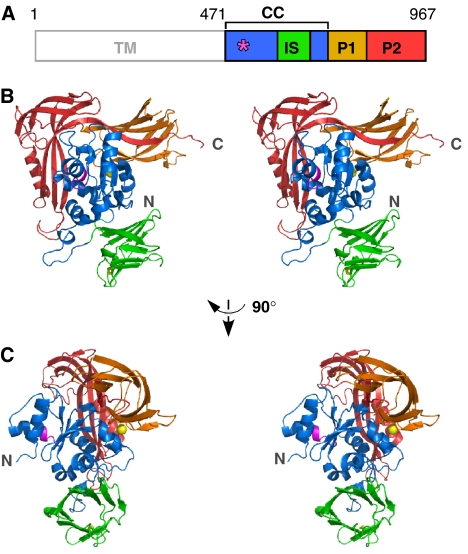
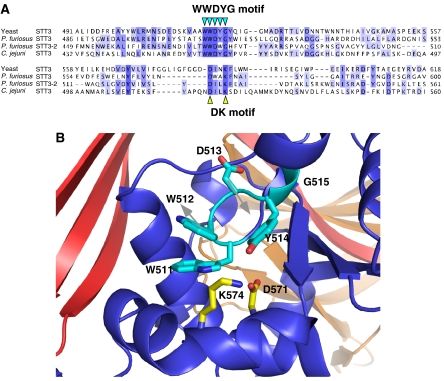
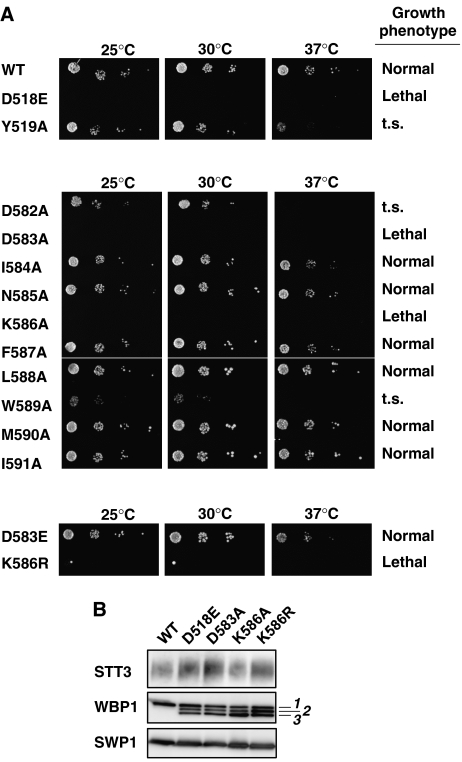
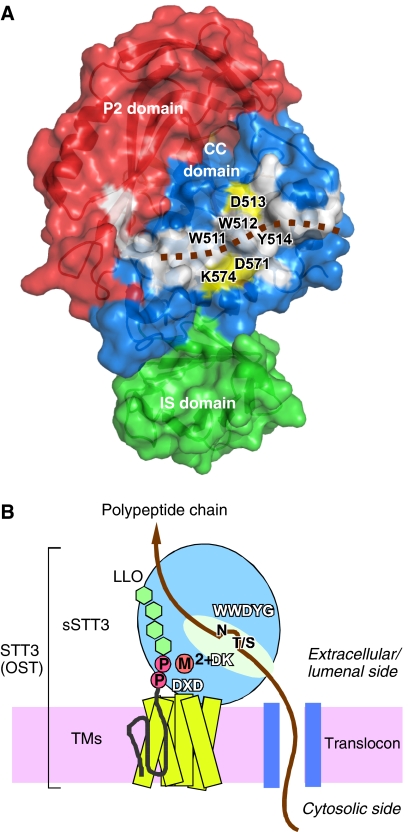
Asn-glycosylation is widespread not only in eukaryotes but also in archaea and some eubacteria. Oligosaccharyltransferase (OST) catalyzes the co-translational transfer of an oligosaccharide from a lipid donor to an asparagine residue in nascent polypeptide chains. Here, we report that a thermophilic archaeon, Pyrococcus furiosus OST is composed of the STT3 protein alone, and catalyzes the transfer of a heptasaccharide, containing one hexouronate and two pentose residues, onto peptides in an Asn-X-Thr/Ser-motif-dependent manner. We also determined the 2.7-A resolution crystal structure of the C-terminal soluble domain of Pyrococcus STT3. The structure-based multiple sequence alignment revealed a new motif, DxxK, which is adjacent to the well-conserved WWDYG motif in the tertiary structure. The mutagenesis of the DK motif residues in yeast STT3 revealed the essential role of the motif in the catalytic activity. The function of this motif may be related to the binding of the pyrophosphate group of lipid-linked oligosaccharide donors through a transiently bound cation. Our structure provides the first structural insights into the formation of the oligosaccharide-asparagine bond.
Figures
References
-
- Brünger AT, Adams PD, Clore GM, DeLano WL, Gros P, Grosse-Kunstleve RW, Jiang JS, Kuszewski J, Nilges M, Pannu NS, Read RJ, Rice LM, Simonson T, Warren GL (1998) Crystallography & NMR system: a new software suite for macromolecular structure determination. Acta Crystallogr D Biol Crystallogr 54: 905–921 - PubMed
-
- Burda P, Aebi M (1999) The dolichol pathway of N-linked glycosylation. Biochim Biophys Acta 1426: 239–257 - PubMed
-
- Chaban B, Voisin S, Kelly J, Logan SM, Jarrell KF (2006) Identification of genes involved in the biosynthesis and attachment of Methanococcus voltae N-linked glycans: insight into N-linked glycosylation pathways in Archaea. Mol Microbiol 61: 259–268 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
