

Predicting preferential DNA vector insertion sites: implications for functional genomics and gene therapy
- PMID: 18047689
- PMCID: PMC2106846
- DOI: 10.1186/gb-2007-8-s1-s12
Predicting preferential DNA vector insertion sites: implications for functional genomics and gene therapy
Abstract
Viral and transposon vectors have been employed in gene therapy as well as functional genomics studies. However, the goals of gene therapy and functional genomics are entirely different; gene therapists hope to avoid altering endogenous gene expression (especially the activation of oncogenes), whereas geneticists do want to alter expression of chromosomal genes. The odds of either outcome depend on a vector's preference to integrate into genes or control regions, and these preferences vary between vectors. Here we discuss the relative strengths of DNA vectors over viral vectors, and review methods to overcome barriers to delivery inherent to DNA vectors. We also review the tendencies of several classes of retroviral and transposon vectors to target DNA sequences, genes, and genetic elements with respect to the balance between insertion preferences and oncogenic selection. Theoretically, knowing the variables that affect integration for various vectors will allow researchers to choose the vector with the most utility for their specific purposes. The three principle benefits from elucidating factors that affect preferences in integration are as follows: in gene therapy, it allows assessment of the overall risks for activating an oncogene or inactivating a tumor suppressor gene that could lead to severe adverse effects years after treatment; in genomic studies, it allows one to discern random from selected integration events; and in gene therapy as well as functional genomics, it facilitates design of vectors that are better targeted to specific sequences, which would be a significant advance in the art of transgenesis.
Figures
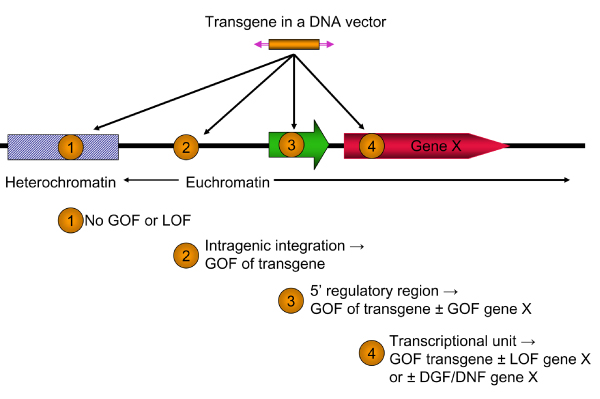
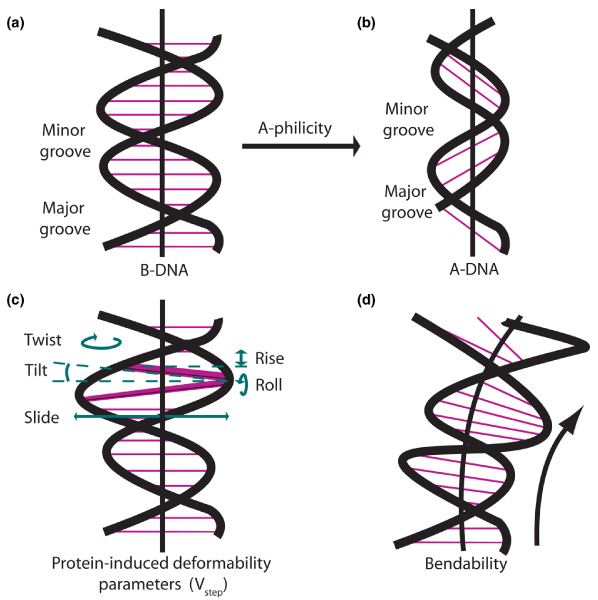
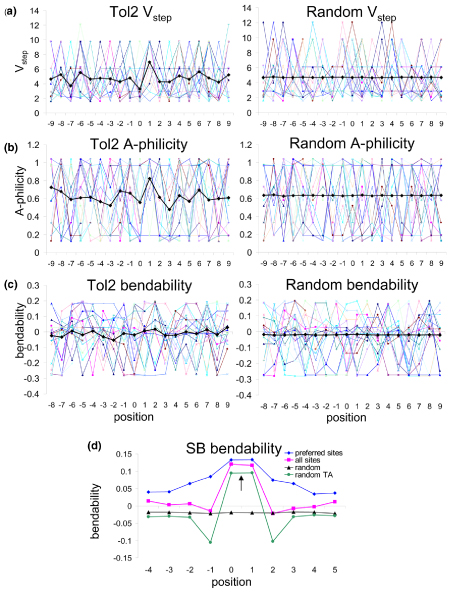
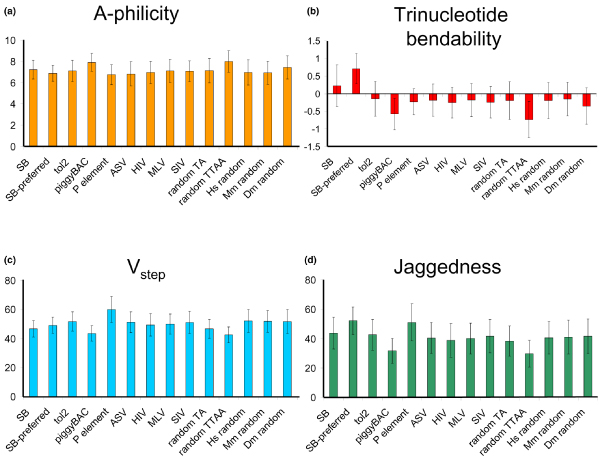
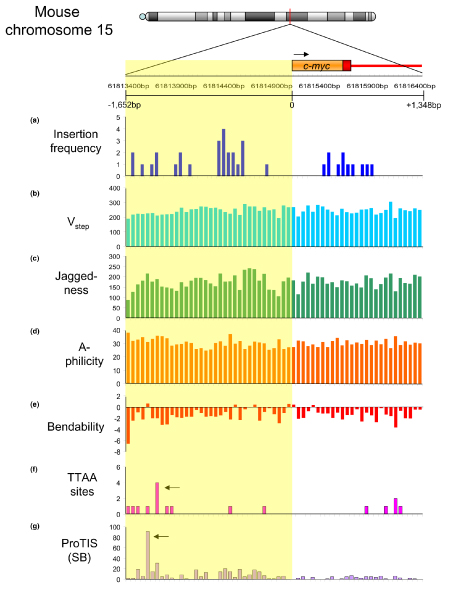
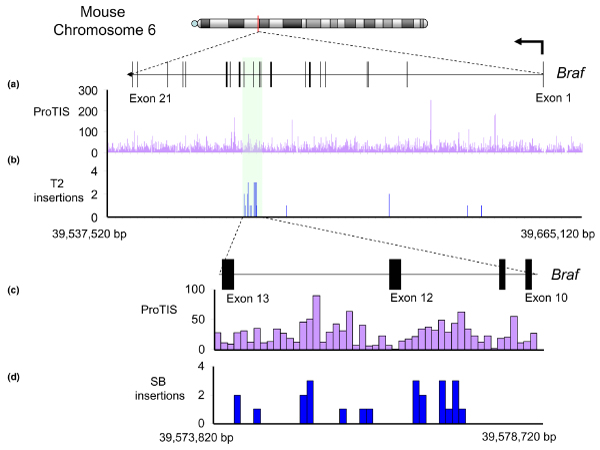
References
-
- Mikkers H, Berns A. Retroviral insertional mutagenesis: tagging cancer pathways. Adv Cancer Res. 2003;88:53–99. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
