

Filling the gap between identified neuroblasts and neurons in crustaceans adds new support for Tetraconata
- PMID: 18048285
- PMCID: PMC2596827
- DOI: 10.1098/rspb.2007.1391
Filling the gap between identified neuroblasts and neurons in crustaceans adds new support for Tetraconata
Abstract
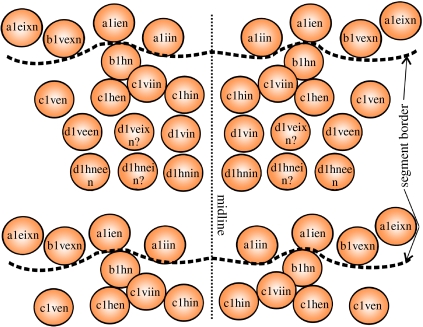
The complex spatio-temporal patterns of development and anatomy of nervous systems play a key role in our understanding of arthropod evolution. However, the degree of resolution of neural processes is not always detailed enough to claim homology between arthropod groups. One example is neural precursors and their progeny in crustaceans and insects. Pioneer neurons of crustaceans and insects show some similarities that indicate homology. In contrast, the differentiation of insect and crustacean neuroblasts (NBs) shows profound differences and their homology is controversial. For Drosophila and grasshoppers, the complete lineage of several NBs up to formation of pioneer neurons is known. Apart from data on median NBs no comparable results exist for Crustacea. Accordingly, it is not clear where the crustacean pioneer neurons come from and whether there are NBs lateral to the midline homologous to those of insects. To fill this gap, individual NBs in the ventral neuroectoderm of the crustacean Orchestia cavimana were labelled in vivo with a fluorescent dye. A partial neuroblast map was established and for the first time lineages from individual NBs to identified pioneer neurons were established in a crustacean. Our data strongly suggest homology of NBs and their lineages, providing further evidence for a close insect-crustacean relationship.
Figures





References
-
- Bate C.M. Embryogenesis of an insect nervous system I. A map of the thoracic and abdominal neuroblasts in Locusta migratoria. J. Embryol. Exp. Morphol. 1976;35:107–123. - PubMed
-
- Boore J.L, Lavrov D.V, Brown W.M. Gene translocation links insects and crustaceans. Nature. 1998;392:667–668. doi:10.1038/33577 - DOI - PubMed
-
- Bossing T, Udolph G, Doe C.Q, Technau G.M. The embryonic central nervous system lineages of Drosophila melanogaster. I. Neuroblast lineages derived from the ventral half of the neurectoderm. Dev. Biol. 1996;179:41–64. doi:10.1006/dbio.1996.0240 - DOI - PubMed
-
- Broadus J, Doe C.Q. Evolution of neuroblast identity: seven-up and prospero expression reveal homologous and divergent neuroblast fates in Drosophila and Schistocerca. Development. 1995;121:3989–3996. - PubMed
-
- Broadus J, Skeath J.B, Spana E.P, Bossing T, Technau G.M, Doe C.Q. New neuroblast markers and the origin of the aCC/pCC neurons in the Drosophila central nervous system. Mech. Dev. 1995;53:393–402. doi:10.1016/0925-4773(95)00454-8 - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous