

Individual variation and the endocrine regulation of behaviour and physiology in birds: a cellular/molecular perspective
- PMID: 18048288
- PMCID: PMC2606728
- DOI: 10.1098/rstb.2007.0010
Individual variation and the endocrine regulation of behaviour and physiology in birds: a cellular/molecular perspective
Abstract
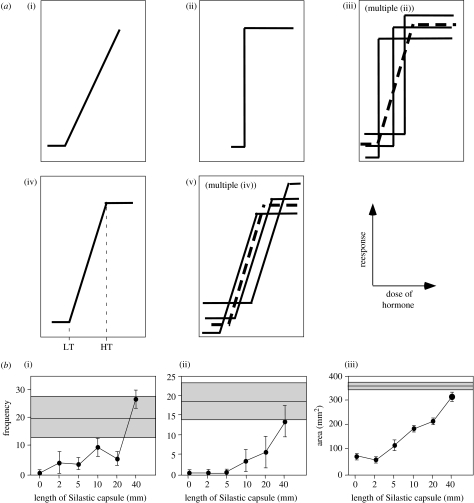
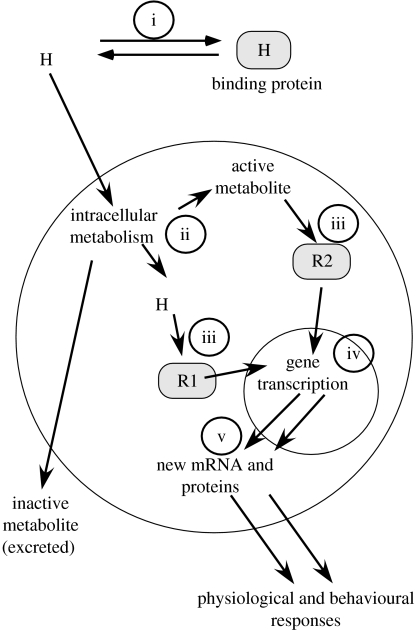
Investigations of the cellular and molecular mechanisms of physiology and behaviour have generally avoided attempts to explain individual differences. The goal has rather been to discover general processes. However, understanding the causes of individual variation in many phenomena of interest to avian eco-physiologists will require a consideration of such mechanisms. For example, in birds, changes in plasma concentrations of steroid hormones are important in the activation of social behaviours related to reproduction and aggression. Attempts to explain individual variation in these behaviours as a function of variation in plasma hormone concentrations have generally failed. Cellular variables related to the effectiveness of steroid hormone have been useful in some cases. Steroid hormone target sensitivity can be affected by variables such as metabolizing enzyme activity, hormone receptor expression as well as receptor cofactor expression. At present, no general theory has emerged that might provide a clear guidance when trying to explain individual variability in birds or in any other group of vertebrates. One strategy is to learn from studies of large units of intraspecific variation such as population or sex differences to provide ideas about variables that might be important in explaining individual variation. This approach along with the use of newly developed molecular genetic tools represents a promising avenue for avian eco-physiologists to pursue.
Figures
References
-
- Absil P, Braquenier J.B, Balthazart J, Ball G.F. Effects of lesions of nucleus taeniae on appetitive and consummatory aspects of male sexual behavior in Japanese quail. Brain Behav. Evol. 2002;60:13–35. doi:10.1159/000064119 - DOI - PubMed
-
- Adkins E.K. Hormonal basis of sexual differentiation in the Japanese quail. J. Comp. Physiol. Psychol. 1975;89:61–71. doi:10.1037/h0076406 - DOI - PubMed
-
- Adkins-Regan E. Princeton University Press; Princeton, NJ: 2005. Hormones and animal social behavior.
-
- Arnold A.P. The effects of castration and androgen replacement on song courtship, and aggression in zebra finches (Poephila guttata) J. Exp. Zool. 1975;191:309–326. doi:10.1002/jez.1401910302 - DOI - PubMed
-
- Arnold A.P. Developmental plasticity in neural circuits controlling birdsong: sexual differentiation and the neural basis of learning. J. Neurobiol. 1992;23:1506–1528. doi:10.1002/neu.480231010 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources