

A helix-breaking mutation in TRPML3 leads to constitutive activity underlying deafness in the varitint-waddler mouse
- PMID: 18048323
- PMCID: PMC2148332
- DOI: 10.1073/pnas.0709846104
A helix-breaking mutation in TRPML3 leads to constitutive activity underlying deafness in the varitint-waddler mouse
Abstract
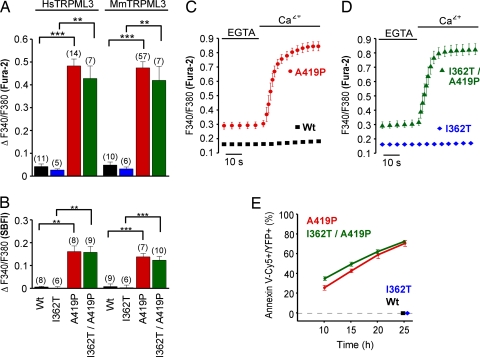
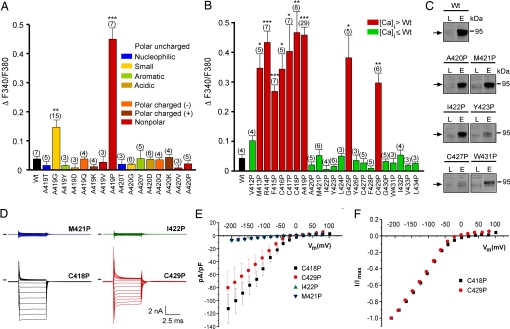
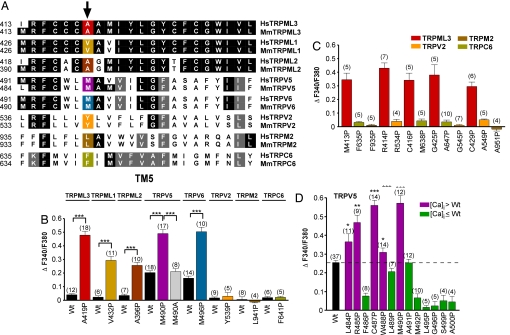
Homozygote varitint-waddler (Va) mice, expressing a mutant isoform (A419P) of TRPML3 (mucolipin 3), are profoundly deaf and display vestibular and pigmentation deficiencies, sterility, and perinatal lethality. Here we show that the varitint-waddler isoform of TRPML3 carrying an A419P mutation represents a constitutively active cation channel that can also be identified in native varitint-waddler hair cells as a distinct inwardly rectifying current. We hypothesize that the constitutive activation of TRPML3 occurs as a result of a helix-breaking proline substitution in transmembrane-spanning domain 5 (TM5). A proline substitution scan demonstrated that the inner third of TRPML3's TM5 is highly susceptible to proline-based kinks. Proline substitutions in TM5 of other TRP channels revealed that TRPML1, TRPML2, TRPV5, and TRPV6 display a similar susceptibility at comparable positions, whereas other TRP channels were not affected. We conclude that the molecular basis for deafness in the varitint-waddler mouse is the result of hair cell death caused by constitutive TRPML3 activity. To our knowledge, our study provides the first direct mechanistic link of a mutation in a TRP ion channel with mammalian hearing loss.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
