

Vitamin B1 biosynthesis in plants requires the essential iron sulfur cluster protein, THIC
- PMID: 18048325
- PMCID: PMC2148341
- DOI: 10.1073/pnas.0709597104
Vitamin B1 biosynthesis in plants requires the essential iron sulfur cluster protein, THIC
Abstract
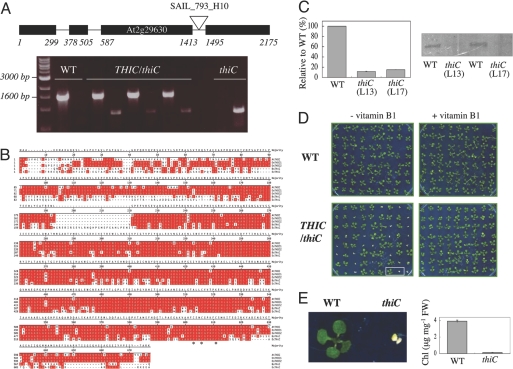
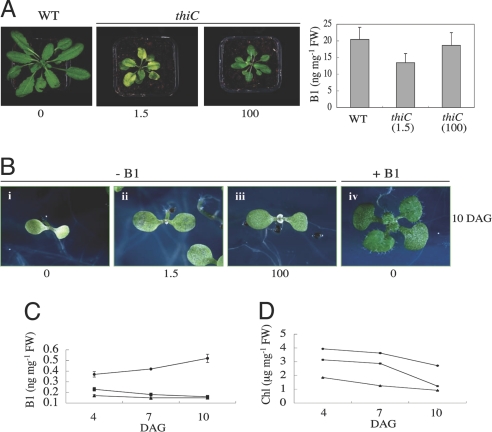
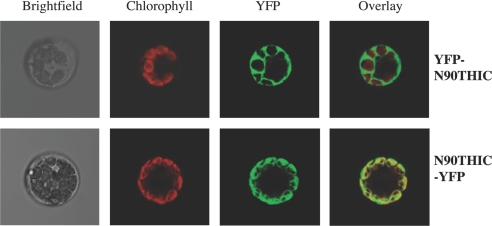
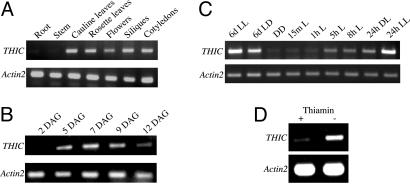
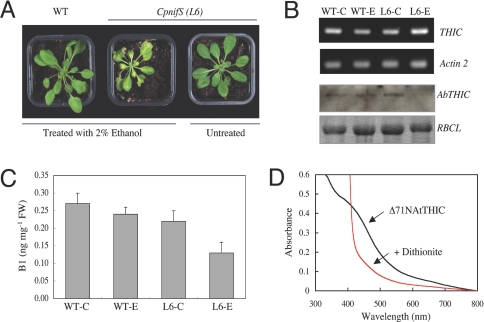
Vitamin B1 (thiamin) is an essential compound in all organisms acting as a cofactor in key metabolic reactions and has furthermore been implicated in responses to DNA damage and pathogen attack in plants. Despite the fact that it was discovered almost a century ago and deficiency is a widespread health problem, much remains to be deciphered about its biosynthesis. The vitamin is composed of a thiazole and pyrimidine heterocycle, which can be synthesized by prokaryotes, fungi, and plants. Plants are the major source of the vitamin in the human diet, yet little is known about the biosynthesis of the compound therein. In particular, it has never been verified whether the pyrimidine heterocycle is derived from purine biosynthesis through the action of the THIC protein as in bacteria, rather than vitamin B6 and histidine as demonstrated for fungi. Here, we identify a homolog of THIC in Arabidopsis and demonstrate its essentiality not only for vitamin B1 biosynthesis, but also plant viability. This step takes place in the chloroplast and appears to be regulated at several levels, including through the presence of a riboswitch in the 3'-untranslated region of THIC. Strong evidence is provided for the involvement of an iron-sulfur cluster in the remarkable chemical rearrangement reaction catalyzed by the THIC protein for which there is no chemical precedent. The results suggest that vitamin B1 biosynthesis in plants is in fact more similar to prokaryotic counterparts and that the THIC protein is likely to be the key regulatory protein in the pathway.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
