

Regulation of Akt/PKB activity by P21-activated kinase in cardiomyocytes
- PMID: 18054038
- PMCID: PMC2278035
- DOI: 10.1016/j.yjmcc.2007.10.016
Regulation of Akt/PKB activity by P21-activated kinase in cardiomyocytes
Abstract
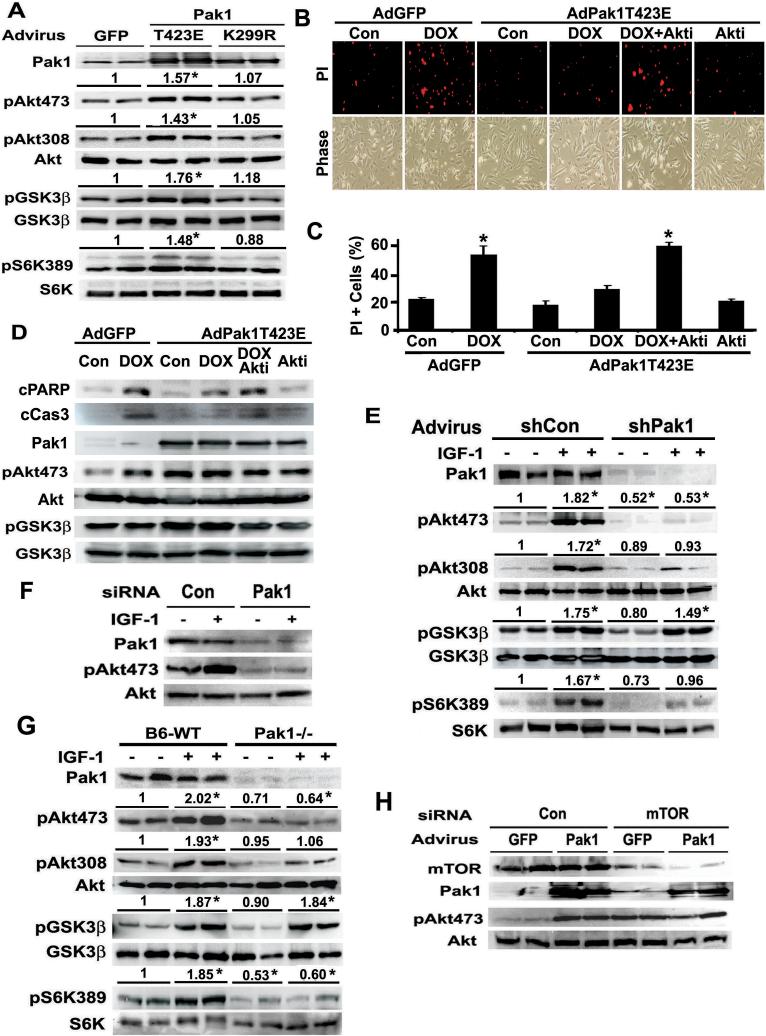
Akt/PKB is a critical regulator of cardiac function and morphology, and its activity is governed by dual phosphorylation at active loop (Thr308) by phosphoinositide-dependent protein kinase-1 (PDK1) and at carboxyl-terminal hydrophobic motif (Ser473) by a putative PDK2. P21-activated kinase-1 (Pak1) is a serine/threonine protein kinase implicated in the regulation of cardiac hypertrophy and contractility and was shown previously to activate Akt through an undefined mechanism. Here we report Pak1 as a potential PDK2 that is essential for Akt activity in cardiomyocytes. Both Pak1 and Akt can be activated by multiple hypertrophic stimuli or growth factors in a phosphatidylinositol-3-kinase (PI3K)-dependent manner. Pak1 overexpression induces Akt phosphorylation at both Ser473 and Thr308 in cardiomyocytes. Conversely, silencing or inactivating Pak1 gene diminishes Akt phosphorylation in vitro and in vivo. Purified Pak1 can directly phosphorylate Akt only at Ser473, suggesting that Pak1 may be a relevant PDK2 responsible for AKT Ser473 phosphorylation in cardiomyocytes. In addition, Pak1 protects cardiomyocytes from cell death, which is blocked by Akt inhibition. Our results connect two important regulators of cellular physiological functions and provide a potential mechanism for Pak1 signaling in cardiomyocytes.
Figures


Similar articles
-
PDK1 acquires PDK2 activity in the presence of a synthetic peptide derived from the carboxyl terminus of PRK2.Curr Biol. 1999 Apr 22;9(8):393-404. doi: 10.1016/s0960-9822(99)80186-9. Curr Biol. 1999. PMID: 10226025
-
Association of PI3K-Akt signaling pathway with digitalis-induced hypertrophy of cardiac myocytes.Am J Physiol Cell Physiol. 2007 Nov;293(5):C1489-97. doi: 10.1152/ajpcell.00158.2007. Epub 2007 Aug 29. Am J Physiol Cell Physiol. 2007. PMID: 17728397
-
Interleukin-18 is a pro-hypertrophic cytokine that acts through a phosphatidylinositol 3-kinase-phosphoinositide-dependent kinase-1-Akt-GATA4 signaling pathway in cardiomyocytes.J Biol Chem. 2005 Feb 11;280(6):4553-67. doi: 10.1074/jbc.M411787200. Epub 2004 Dec 1. J Biol Chem. 2005. PMID: 15574430
-
PIKKing on PKB: regulation of PKB activity by phosphorylation.Curr Opin Cell Biol. 2009 Apr;21(2):256-61. doi: 10.1016/j.ceb.2009.02.002. Epub 2009 Mar 19. Curr Opin Cell Biol. 2009. PMID: 19303758 Review.
-
AKT signalling in the failing heart.Eur J Heart Fail. 2011 Aug;13(8):825-9. doi: 10.1093/eurjhf/hfr080. Epub 2011 Jun 30. Eur J Heart Fail. 2011. PMID: 21724622 Free PMC article. Review.
Cited by
-
Genetic deletion of trkB.T1 increases neuromuscular function.Am J Physiol Cell Physiol. 2012 Jan 1;302(1):C141-53. doi: 10.1152/ajpcell.00469.2010. Epub 2011 Oct 5. Am J Physiol Cell Physiol. 2012. PMID: 21865582 Free PMC article.
-
The mTORC2 signaling network: targets and cross-talks.Biochem J. 2024 Jan 25;481(2):45-91. doi: 10.1042/BCJ20220325. Biochem J. 2024. PMID: 38270460 Free PMC article. Review.
-
Endoplasmic reticulum stress causes insulin resistance by inhibiting delivery of newly synthesized insulin receptors to the cell surface.Mol Biol Cell. 2020 Nov 1;31(23):2597-2629. doi: 10.1091/mbc.E18-01-0013. Epub 2020 Sep 2. Mol Biol Cell. 2020. PMID: 32877278 Free PMC article.
-
Cardiac deficiency of P21-activated kinase 1 promotes atrial arrhythmogenesis in mice following adrenergic challenge.Philos Trans R Soc Lond B Biol Sci. 2023 Jun 19;378(1879):20220168. doi: 10.1098/rstb.2022.0168. Epub 2023 May 1. Philos Trans R Soc Lond B Biol Sci. 2023. PMID: 37122217 Free PMC article.
-
Bta-miR-106b Regulates Bovine Mammary Epithelial Cell Proliferation, Cell Cycle, and Milk Protein Synthesis by Targeting the CDKN1A Gene.Genes (Basel). 2022 Dec 7;13(12):2308. doi: 10.3390/genes13122308. Genes (Basel). 2022. PMID: 36553575 Free PMC article.
References
-
- Shiojima I, Walsh K. Regulation of cardiac growth and coronary angiogenesis by the Akt/PKB signaling pathway. Genes Dev. 2006;20:3347–3365. - PubMed
-
- Alessi DR, James SR, Downes CP, Holmes AB, Gaffney PR, Reese CB, Cohen P. Characterization of a 3-phosphoinositide-dependent protein kinase which phosphorylates and activates protein kinase Balpha. Curr Biol. 1997;7:261–269. - PubMed
-
- Dong LQ, Liu F. PDK2: the missing piece in the receptor tyrosine kinase signaling pathway puzzle. Am J Physiol Endocrinol Metab. 2005;289:E187–196. - PubMed
-
- Guertin DA, Stevens DM, Thoreen CC, Burds AA, Kalaany NY, Moffat J, Brown M, Fitzgerald KJ, Sabatini DM. Ablation in mice of the mTORC components raptor, rictor, or mLST8 reveals that mTORC2 is required for signaling to Akt-FOXO and PKCalpha, but not S6K1. Dev Cell. 2006;11:859–871. - PubMed
-
- Bokoch GM. Biology of the p21-activated kinases. Annu Rev Biochem. 2003;72:743–781. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
