

Gating of Sema3E/PlexinD1 signaling by neuropilin-1 switches axonal repulsion to attraction during brain development
- PMID: 18054858
- PMCID: PMC2700040
- DOI: 10.1016/j.neuron.2007.10.019
Gating of Sema3E/PlexinD1 signaling by neuropilin-1 switches axonal repulsion to attraction during brain development
Abstract
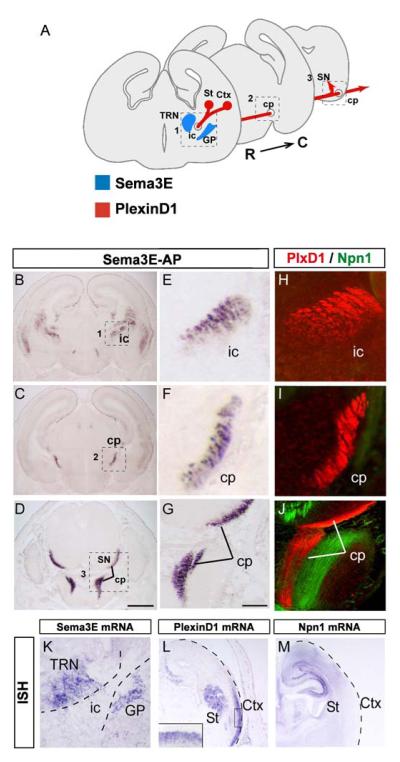
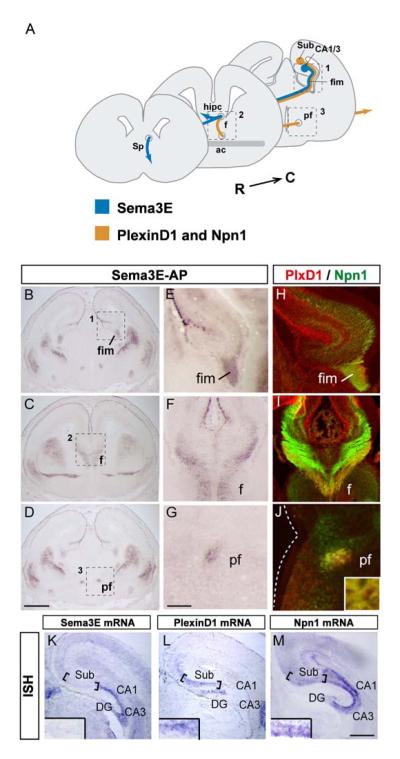
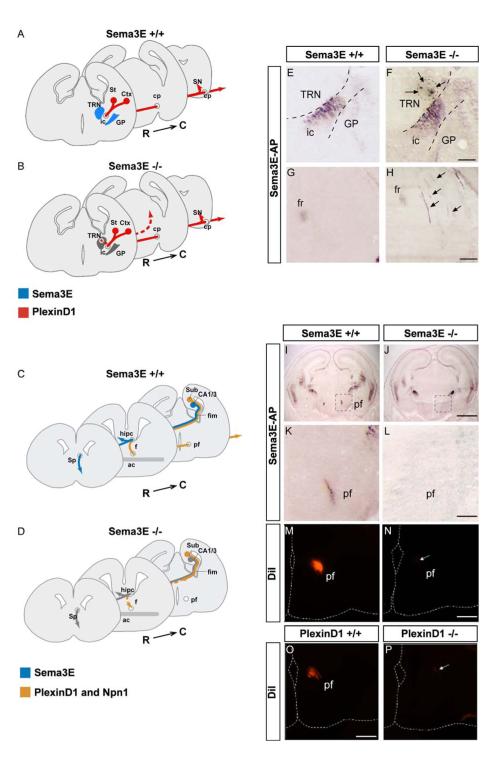
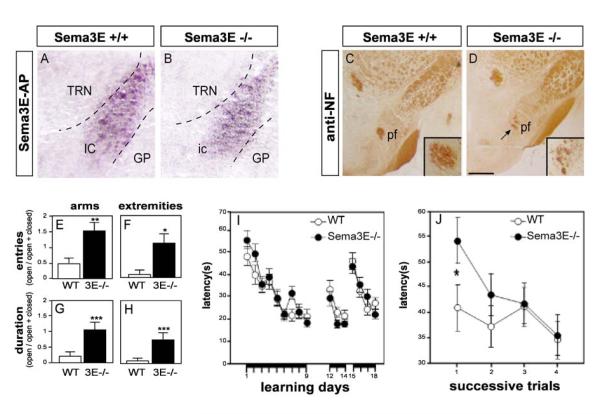
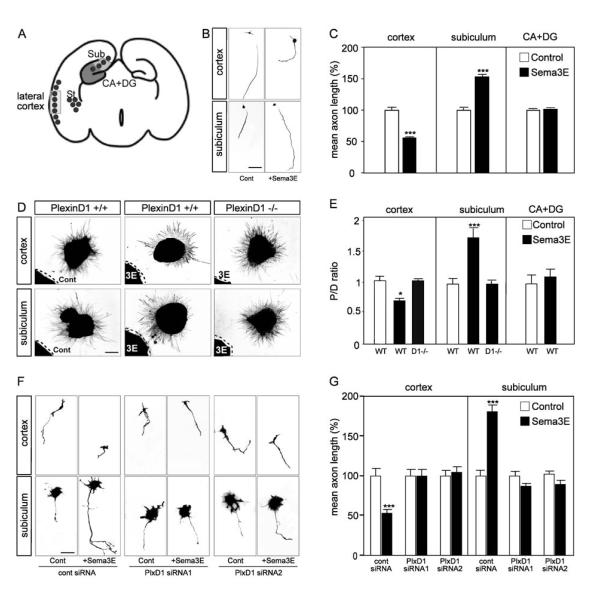
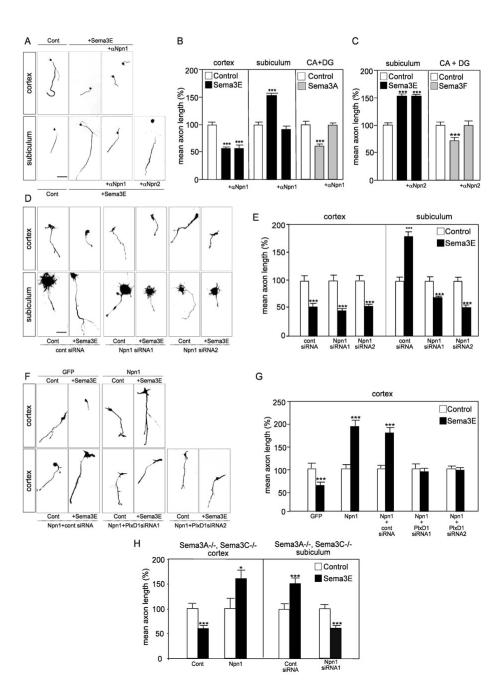
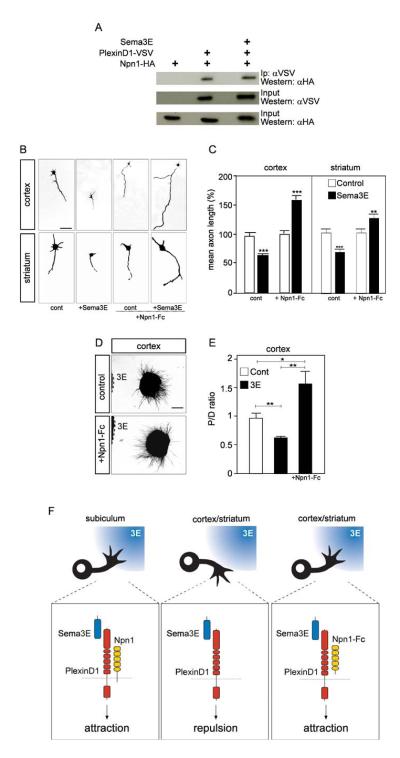
The establishment of functional neural circuits requires the guidance of axons in response to the actions of secreted and cell-surface molecules such as the semaphorins. Semaphorin 3E and its receptor PlexinD1 are expressed in the brain, but their functions are unknown. Here, we show that Sema3E/PlexinD1 signaling plays an important role in initial development of descending axon tracts in the forebrain. Early errors in axonal projections are reflected in behavioral deficits in Sema3E null mutant mice. Two distinct signaling mechanisms can be distinguished downstream of Sema3E. On corticofugal and striatonigral neurons expressing PlexinD1 but not Neuropilin-1, Sema3E acts as a repellent. In contrast, on subiculo-mammillary neurons coexpressing PlexinD1 and Neuropilin-1, Sema3E acts as an attractant. The extracellular domain of Neuropilin-1 is sufficient to convert repulsive signaling by PlexinD1 to attraction. Our data therefore reveal a "gating" function of neuropilins in semaphorin-plexin signaling during the assembly of forebrain neuronal circuits.
Figures
Similar articles
-
Semaphorin 3d and semaphorin 3e direct endothelial motility through distinct molecular signaling pathways.J Biol Chem. 2014 Jun 27;289(26):17971-9. doi: 10.1074/jbc.M113.544833. Epub 2014 May 13. J Biol Chem. 2014. PMID: 24825896 Free PMC article.
-
Plexin-a4 mediates axon-repulsive activities of both secreted and transmembrane semaphorins and plays roles in nerve fiber guidance.J Neurosci. 2005 Apr 6;25(14):3628-37. doi: 10.1523/JNEUROSCI.4480-04.2005. J Neurosci. 2005. PMID: 15814794 Free PMC article.
-
Specificity of monosynaptic sensory-motor connections imposed by repellent Sema3E-PlexinD1 signaling.Cell Rep. 2013 Nov 14;5(3):748-58. doi: 10.1016/j.celrep.2013.10.005. Epub 2013 Nov 7. Cell Rep. 2013. PMID: 24210822 Free PMC article.
-
Semaphorins and their receptors in olfactory axon guidance.Cell Mol Biol (Noisy-le-grand). 1999 Sep;45(6):763-79. Cell Mol Biol (Noisy-le-grand). 1999. PMID: 10541474 Review.
-
Knowing how to navigate: mechanisms of semaphorin signaling in the nervous system.Sci STKE. 2002 Feb 12;2002(119):re1. doi: 10.1126/stke.2002.119.re1. Sci STKE. 2002. PMID: 11842242 Review.
Cited by
-
Cortical RORβ is required for layer 4 transcriptional identity and barrel integrity.Elife. 2020 Aug 27;9:e52370. doi: 10.7554/eLife.52370. Elife. 2020. PMID: 32851975 Free PMC article.
-
Molecular parallels between neural and vascular development.Cold Spring Harb Perspect Med. 2013 Jan 1;3(1):a006551. doi: 10.1101/cshperspect.a006551. Cold Spring Harb Perspect Med. 2013. PMID: 23024177 Free PMC article. Review.
-
Contribution of semaphorins to the formation of the peripheral nervous system in higher vertebrates.Cell Adh Migr. 2016 Nov;10(6):593-603. doi: 10.1080/19336918.2016.1243644. Epub 2016 Oct 7. Cell Adh Migr. 2016. PMID: 27715392 Free PMC article. Review.
-
Guidance from above: common cues direct distinct signaling outcomes in vascular and neural patterning.Trends Cell Biol. 2009 Mar;19(3):99-110. doi: 10.1016/j.tcb.2009.01.001. Epub 2009 Feb 4. Trends Cell Biol. 2009. PMID: 19200729 Free PMC article. Review.
-
GIPC proteins negatively modulate Plexind1 signaling during vascular development.Elife. 2019 May 3;8:e30454. doi: 10.7554/eLife.30454. Elife. 2019. PMID: 31050647 Free PMC article.
References
-
- Bachelder RE, Lipscomb EA, Lin X, Wendt MA, Chadborn NH, Eickholt BJ, Mercurio AM. Competing autocrine pathways involving alternative neuropilin-1 ligands regulate chemotaxis of carcinoma cells. Cancer Res. 2003;63:5230–5233. - PubMed
-
- Bagnard D, Lohrum M, Uziel D, Puschel AW, Bolz J. Semaphorins act as attractive and repulsive guidance signals during the development of cortical projections. Development. 1998;125:5043–5053. - PubMed
-
- Bagri A, Tessier-Lavigne M. Neuropilins as Semaphorin receptors: in vivo functions in neuronal cell migration and axon guidance. Adv Exp Med Biol. 2002;515:13–31. - PubMed
-
- Bagri A, Marin O, Plump AS, Mak J, Pleasure SJ, Rubenstein JL, Tessier-Lavigne M. Slit proteins prevent midline crossing and determine the dorsoventral position of major axonal pathways in the mammalian forebrain. Neuron. 2002;33:233–248. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
