

Synthesis of the D2 protein of photosystem II in Chlamydomonas is controlled by a high molecular mass complex containing the RNA stabilization factor Nac2 and the translational activator RBP40
- PMID: 18055611
- PMCID: PMC2174876
- DOI: 10.1105/tpc.107.051722
Synthesis of the D2 protein of photosystem II in Chlamydomonas is controlled by a high molecular mass complex containing the RNA stabilization factor Nac2 and the translational activator RBP40
Abstract
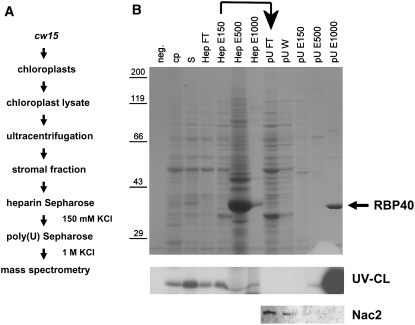
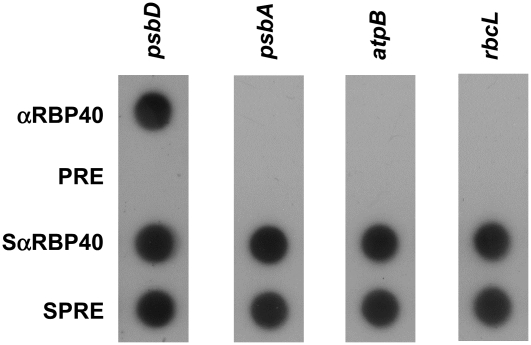
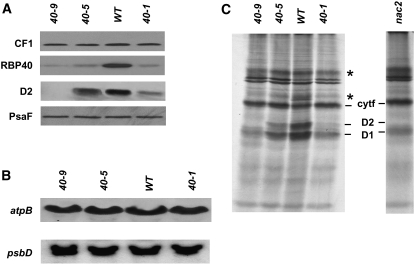
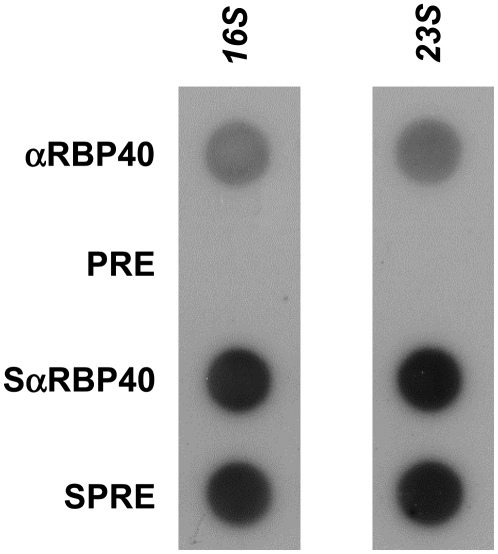
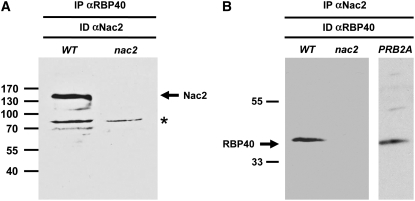
Gene expression in chloroplasts is regulated mainly at the posttranscriptional level. In the green alga Chlamydomonas reinhardtii, synthesis of the D2 protein (PsbD), which is the rate-determining subunit for the assembly of photosystem II, depends on the RNA stability factor Nac2. In addition, the RNA binding protein RBP40 has been implicated in translational control via a U-rich element in the 5' untranslated region (5'UTR) of the psbD mRNA. Here, we report the identification of the RBP40 gene based on mass spectrometric analysis of its purified product. Unexpectedly, this was found to be identical to the previously described RNA binding protein RB38, which had been suggested to be involved in the regulation of D1 protein synthesis. However, we show that RBP40 binds to the psbD 5'UTR in a Nac2-dependent fashion both in vitro and in vivo. Molecular characterization of RBP40 RNA interference lines confirmed that RBP40 specifically affects the initiation of D2 synthesis. Native polyacrylamide gel electrophoresis, coimmunoprecipitation, and sedimentation analyses revealed that Nac2 and RBP40 form parts of a complex of 550 kD that is displaced from the psbD mRNA prior to polysome assembly. Together, these data indicate that the processes of 5'UTR-mediated RNA stabilization and translation initiation are tightly coupled in Chlamydomonas.
Figures





Similar articles
-
An intermolecular disulfide-based light switch for chloroplast psbD gene expression in Chlamydomonas reinhardtii.Plant J. 2012 Nov;72(3):378-89. doi: 10.1111/j.1365-313X.2012.05083.x. Epub 2012 Sep 13. Plant J. 2012. PMID: 22725132
-
The Nac2 gene of Chlamydomonas encodes a chloroplast TPR-like protein involved in psbD mRNA stability.EMBO J. 2000 Jul 3;19(13):3366-76. doi: 10.1093/emboj/19.13.3366. EMBO J. 2000. PMID: 10880449 Free PMC article.
-
cis- and trans-Acting determinants for translation of psbD mRNA in Chlamydomonas reinhardtii.Mol Cell Biol. 2000 Nov;20(21):8134-42. doi: 10.1128/MCB.20.21.8134-8142.2000. Mol Cell Biol. 2000. PMID: 11027283 Free PMC article.
-
Loss of chloroplast ClpP elicits an autophagy-like response in Chlamydomonas.Autophagy. 2014 Sep;10(9):1685-6. doi: 10.4161/auto.29960. Epub 2014 Jul 17. Autophagy. 2014. PMID: 25046108 Free PMC article. Review.
-
Novel insights into the function of LHCSR3 in Chlamydomonas reinhardtii.Plant Signal Behav. 2015;10(12):e1058462. doi: 10.1080/15592324.2015.1058462. Plant Signal Behav. 2015. PMID: 26237677 Free PMC article. Review.
Cited by
-
The availability of neither D2 nor CP43 limits the biogenesis of photosystem II in tobacco.Plant Physiol. 2021 Apr 2;185(3):1111-1130. doi: 10.1093/plphys/kiaa052. Plant Physiol. 2021. PMID: 33793892 Free PMC article.
-
Manipulating RuBisCO accumulation in the green alga, Chlamydomonas reinhardtii.Plant Mol Biol. 2011 Jul;76(3-5):397-405. doi: 10.1007/s11103-011-9783-z. Epub 2011 May 24. Plant Mol Biol. 2011. PMID: 21607658
-
Dynamics of Chloroplast Translation during Chloroplast Differentiation in Maize.PLoS Genet. 2016 Jul 14;12(7):e1006106. doi: 10.1371/journal.pgen.1006106. eCollection 2016 Jul. PLoS Genet. 2016. PMID: 27414025 Free PMC article.
-
Function of chloroplast RNA-binding proteins.Cell Mol Life Sci. 2011 Mar;68(5):735-48. doi: 10.1007/s00018-010-0523-3. Epub 2010 Sep 17. Cell Mol Life Sci. 2011. PMID: 20848156 Free PMC article. Review.
-
Micro-algae come of age as a platform for recombinant protein production.Biotechnol Lett. 2010 Oct;32(10):1373-83. doi: 10.1007/s10529-010-0326-5. Epub 2010 Jun 17. Biotechnol Lett. 2010. PMID: 20556634 Free PMC article. Review.
References
-
- Barnes, D., Cohen, A., Bruick, R.K., Kantardjieff, K., Fowler, S., Efuet, E., and Mayfield, S.P. (2004). Identification and characterization of a novel RNA binding protein that associates with the 5′ untranslated region of the chloroplast psbA mRNA. Biochemistry 43 8541–8550. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
