

Motif module map reveals enforcement of aging by continual NF-kappaB activity
- PMID: 18055696
- PMCID: PMC2113026
- DOI: 10.1101/gad.1588507
Motif module map reveals enforcement of aging by continual NF-kappaB activity
Abstract
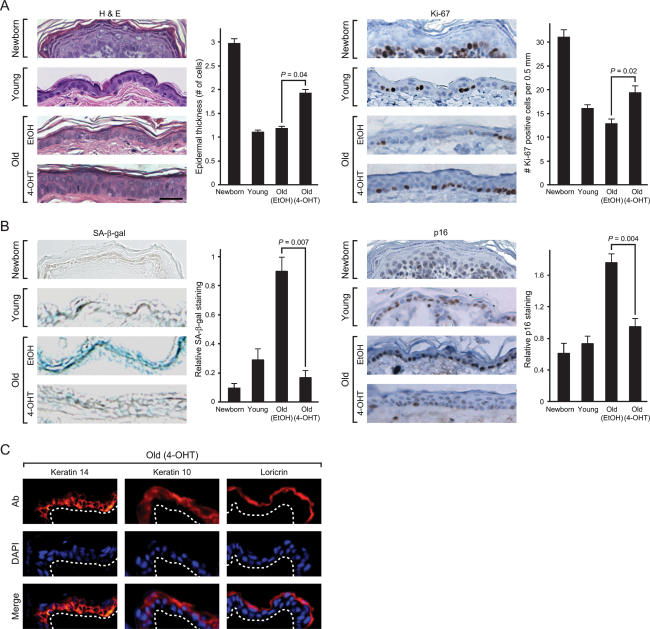
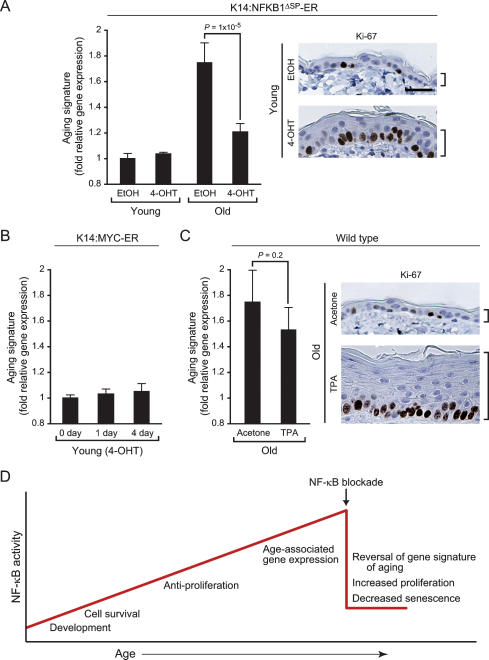
Aging is characterized by specific alterations in gene expression, but their underlying mechanisms and functional consequences are not well understood. Here we develop a systematic approach to identify combinatorial cis-regulatory motifs that drive age-dependent gene expression across different tissues and organisms. Integrated analysis of 365 microarrays spanning nine tissue types predicted fourteen motifs as major regulators of age-dependent gene expression in human and mouse. The motif most strongly associated with aging was that of the transcription factor NF-kappaB. Inducible genetic blockade of NF-kappaB for 2 wk in the epidermis of chronologically aged mice reverted the tissue characteristics and global gene expression programs to those of young mice. Age-specific NF-kappaB blockade and orthogonal cell cycle interventions revealed that NF-kappaB controls cell cycle exit and gene expression signature of aging in parallel but not sequential pathways. These results identify a conserved network of regulatory pathways underlying mammalian aging and show that NF-kappaB is continually required to enforce many features of aging in a tissue-specific manner.
Figures
References
-
- April C.S., Barsh G.S., Barsh G.S. Skin layer-specific transcriptional profiles in normal and recessive yellow (Mc1re/Mc1re) mice. Pigment Cell Res. 2006;19:194–205. - PubMed
-
- Arkan M.C., Hevener A.L., Greten F.R., Maeda S., Li Z.W., Long J.M., Wynshaw-Boris A., Poli G., Olefsky J., Karin M., Hevener A.L., Greten F.R., Maeda S., Li Z.W., Long J.M., Wynshaw-Boris A., Poli G., Olefsky J., Karin M., Greten F.R., Maeda S., Li Z.W., Long J.M., Wynshaw-Boris A., Poli G., Olefsky J., Karin M., Maeda S., Li Z.W., Long J.M., Wynshaw-Boris A., Poli G., Olefsky J., Karin M., Li Z.W., Long J.M., Wynshaw-Boris A., Poli G., Olefsky J., Karin M., Long J.M., Wynshaw-Boris A., Poli G., Olefsky J., Karin M., Wynshaw-Boris A., Poli G., Olefsky J., Karin M., Poli G., Olefsky J., Karin M., Olefsky J., Karin M., Karin M. IKK-β links inflammation to obesity-induced insulin resistance. Nat. Med. 2005;11:191–198. - PubMed
-
- Baur J.A., Pearson K.J., Price N.L., Jamieson H.A., Lerin C., Kalra A., Prabhu V.V., Allard J.S., Lopez-Lluch G., Lewis K., Pearson K.J., Price N.L., Jamieson H.A., Lerin C., Kalra A., Prabhu V.V., Allard J.S., Lopez-Lluch G., Lewis K., Price N.L., Jamieson H.A., Lerin C., Kalra A., Prabhu V.V., Allard J.S., Lopez-Lluch G., Lewis K., Jamieson H.A., Lerin C., Kalra A., Prabhu V.V., Allard J.S., Lopez-Lluch G., Lewis K., Lerin C., Kalra A., Prabhu V.V., Allard J.S., Lopez-Lluch G., Lewis K., Kalra A., Prabhu V.V., Allard J.S., Lopez-Lluch G., Lewis K., Prabhu V.V., Allard J.S., Lopez-Lluch G., Lewis K., Allard J.S., Lopez-Lluch G., Lewis K., Lopez-Lluch G., Lewis K., Lewis K., et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature. 2006;444:337–342. - PMC - PubMed
-
- Bernard D., Gosselin K., Monte D., Vercamer C., Bouali F., Pourtier A., Vandenbunder B., Abbadie C., Gosselin K., Monte D., Vercamer C., Bouali F., Pourtier A., Vandenbunder B., Abbadie C., Monte D., Vercamer C., Bouali F., Pourtier A., Vandenbunder B., Abbadie C., Vercamer C., Bouali F., Pourtier A., Vandenbunder B., Abbadie C., Bouali F., Pourtier A., Vandenbunder B., Abbadie C., Pourtier A., Vandenbunder B., Abbadie C., Vandenbunder B., Abbadie C., Abbadie C. Involvement of Rel/nuclear factor-κB transcription factors in keratinocyte senescence. Cancer Res. 2004;64:472–481. - PubMed
-
- Borneman A.R., Leigh-Bell J.A., Yu H., Bertone P., Gerstein M., Snyder M., Leigh-Bell J.A., Yu H., Bertone P., Gerstein M., Snyder M., Yu H., Bertone P., Gerstein M., Snyder M., Bertone P., Gerstein M., Snyder M., Gerstein M., Snyder M., Snyder M. Target hub proteins serve as master regulators of development in yeast. Genes & Dev. 2006;20:435–448. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases