

Atg7-dependent autophagy promotes neuronal health, stress tolerance, and longevity but is dispensable for metamorphosis in Drosophila
- PMID: 18056421
- PMCID: PMC2081972
- DOI: 10.1101/gad.1600707
Atg7-dependent autophagy promotes neuronal health, stress tolerance, and longevity but is dispensable for metamorphosis in Drosophila
Abstract
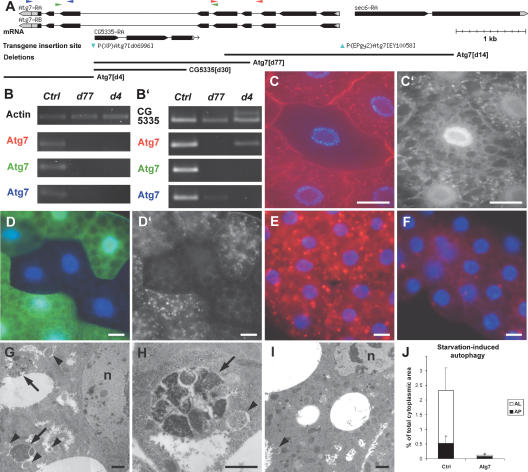
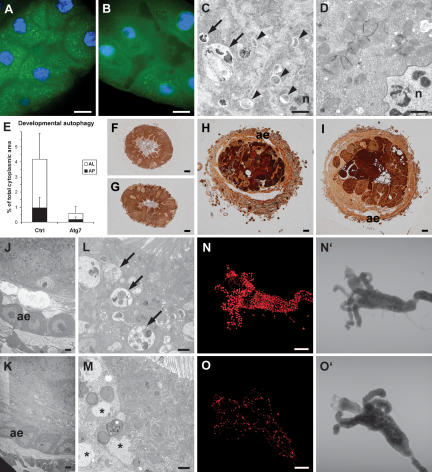
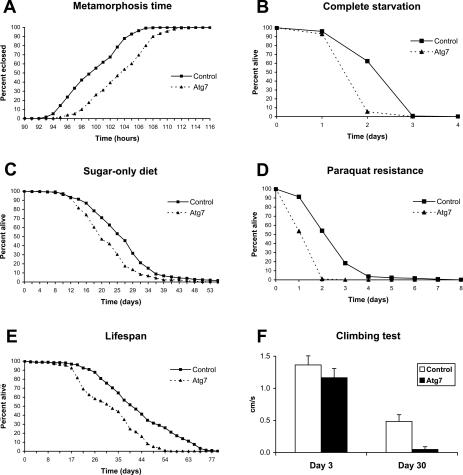
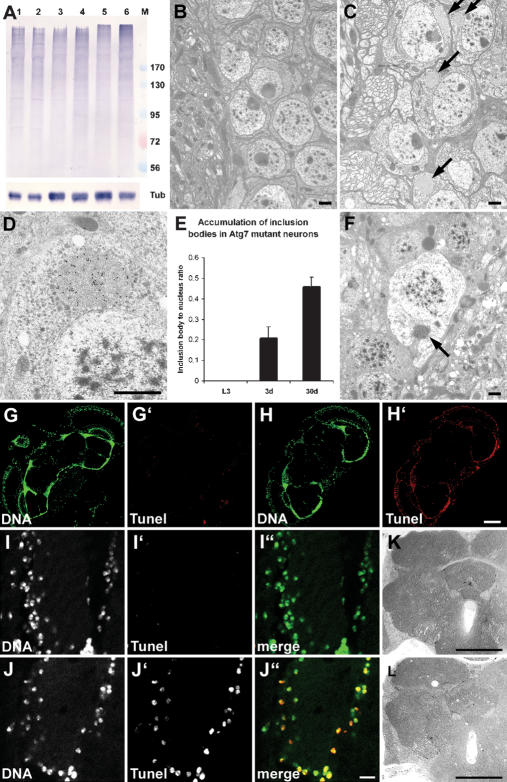
Autophagy, a cellular process of cytoplasmic degradation and recycling, is induced in Drosophila larval tissues during metamorphosis, potentially contributing to their destruction or reorganization. Unexpectedly, we find that flies lacking the core autophagy regulator Atg7 are viable, despite severe defects in autophagy. Although metamorphic cell death is perturbed in Atg7 mutants, the larval-adult midgut transition proceeds normally, with extended pupal development compensating for reduced autophagy. Atg7-/- adults are short-lived, hypersensitive to nutrient and oxidative stress, and accumulate ubiquitin-positive aggregates in degenerating neurons. Thus, normal levels of autophagy are crucial for stress survival and continuous cellular renewal, but not metamorphosis.
Figures
References
-
- Butterworth F.M., Forrest E.C., Forrest E.C. Ultrastructure of the preparative phase of cell death in the larval fat body of Drosophila melanogaster. Tissue Cell. 1984;16:237–250. - PubMed
-
- Cuervo A.M., Bergamini E., Brunk U.T., Droge W., Ffrench M., Terman A., Bergamini E., Brunk U.T., Droge W., Ffrench M., Terman A., Brunk U.T., Droge W., Ffrench M., Terman A., Droge W., Ffrench M., Terman A., Ffrench M., Terman A., Terman A. Autophagy and aging: The importance of maintaining ‘clean’ cells. Autophagy. 2005;1:131–140. - PubMed
-
- Hara T., Nakamura K., Matsui M., Yamamoto A., Nakahara Y., Suzuki-Migishima R., Yokoyama M., Mishima K., Saito I., Okano H., Nakamura K., Matsui M., Yamamoto A., Nakahara Y., Suzuki-Migishima R., Yokoyama M., Mishima K., Saito I., Okano H., Matsui M., Yamamoto A., Nakahara Y., Suzuki-Migishima R., Yokoyama M., Mishima K., Saito I., Okano H., Yamamoto A., Nakahara Y., Suzuki-Migishima R., Yokoyama M., Mishima K., Saito I., Okano H., Nakahara Y., Suzuki-Migishima R., Yokoyama M., Mishima K., Saito I., Okano H., Suzuki-Migishima R., Yokoyama M., Mishima K., Saito I., Okano H., Yokoyama M., Mishima K., Saito I., Okano H., Mishima K., Saito I., Okano H., Saito I., Okano H., Okano H., et al. Suppression of basal autophagy in neural cells causes neurodegenerative disease in mice. Nature. 2006;441:885–889. - PubMed
-
- Klionsky D.J., Cregg J.M., Dunn W.A., Emr S.D., Sakai Y., Sandoval I.V., Sibirny A., Subramani S., Thumm M., Veenhuis M., Cregg J.M., Dunn W.A., Emr S.D., Sakai Y., Sandoval I.V., Sibirny A., Subramani S., Thumm M., Veenhuis M., Dunn W.A., Emr S.D., Sakai Y., Sandoval I.V., Sibirny A., Subramani S., Thumm M., Veenhuis M., Emr S.D., Sakai Y., Sandoval I.V., Sibirny A., Subramani S., Thumm M., Veenhuis M., Sakai Y., Sandoval I.V., Sibirny A., Subramani S., Thumm M., Veenhuis M., Sandoval I.V., Sibirny A., Subramani S., Thumm M., Veenhuis M., Sibirny A., Subramani S., Thumm M., Veenhuis M., Subramani S., Thumm M., Veenhuis M., Thumm M., Veenhuis M., Veenhuis M., et al. A unified nomenclature for yeast autophagy-related genes. Dev. Cell. 2003;5:539–545. - PubMed
-
- Komatsu M., Waguri S., Ueno T., Iwata J., Murata S., Tanida I., Ezaki J., Mizushima N., Ohsumi Y., Uchiyama Y., Waguri S., Ueno T., Iwata J., Murata S., Tanida I., Ezaki J., Mizushima N., Ohsumi Y., Uchiyama Y., Ueno T., Iwata J., Murata S., Tanida I., Ezaki J., Mizushima N., Ohsumi Y., Uchiyama Y., Iwata J., Murata S., Tanida I., Ezaki J., Mizushima N., Ohsumi Y., Uchiyama Y., Murata S., Tanida I., Ezaki J., Mizushima N., Ohsumi Y., Uchiyama Y., Tanida I., Ezaki J., Mizushima N., Ohsumi Y., Uchiyama Y., Ezaki J., Mizushima N., Ohsumi Y., Uchiyama Y., Mizushima N., Ohsumi Y., Uchiyama Y., Ohsumi Y., Uchiyama Y., Uchiyama Y., et al. Impairment of starvation-induced and constitutive autophagy in Atg7-deficient mice. J. Cell Biol. 2005;169:425–434. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases