

Review
doi: 10.1104/pp.107.106641.
New gateways to discovery
Affiliations
- PMID: 18056860
- PMCID: PMC2151732
- DOI: 10.1104/pp.107.106641
Item in Clipboard
Review
New gateways to discovery
Plant Physiol.
2007 Dec.
No abstract available
Figures
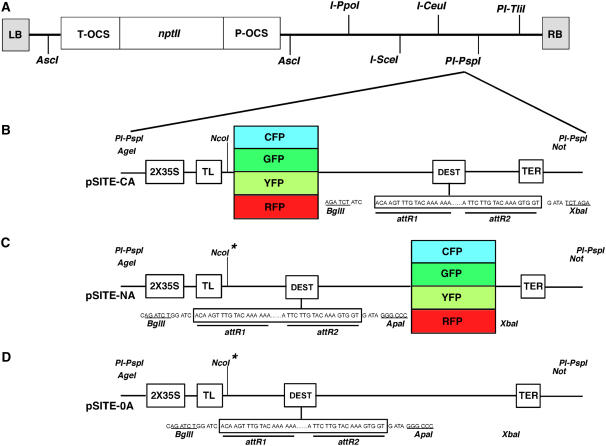
Schematic representations of pSITE vectors. A, All modified pSAT6 cassettes were cloned into the pRCS2-ocs-nptII binary vector at the PI-Psp1 site. The ability to select transgenic plant cells is conferred by the nptII gene, the expression of which is controlled by the ocs promoter (P-OCS) and terminator (T-OCS). B, C-series pSITE vectors for Gateway recombination-mediated construction of binary vectors for expression of proteins of interest fused to the carboxy termini of AFPs. C, N-series pSITE vectors for Gateway recombination-mediated construction of binary vectors for expression of proteins of interest fused to the amino termini of AFPs. D, 0-series pSITE vectors for Gateway recombination-mediated construction of binary vectors for expression of native proteins. Protein expression is controlled by a duplicated CaMV 35S promoter (2X35S) and a tobacco etch virus translational leader (TL). All vectors employ the CaMV35S transcriptional terminator (TER). Nco1*, This restriction site was deleted to create the pSITE-NB and pSITE-0B vectors, thereby allowing translation to initiate at the native start codon on the gene of interest. This figure is a revision of that appearing in Chakrabarty et al. (2007). [See online article for color version of this figure.]
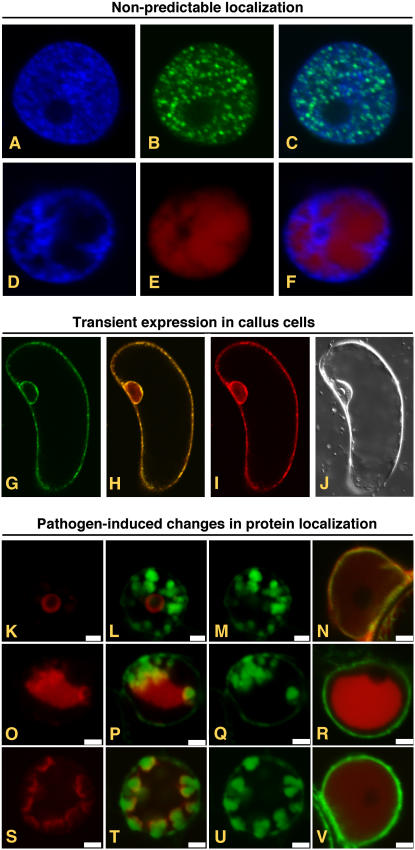
Transient expression of AFPs from pSITE vectors. A to F, Expression of proteins whose subcellular localization cannot be determined in silico. Fluorescence from DAPI (A) and GFP (B) in the nucleus of a cell expressing a GFP:PYDV-P protein. The overlay of A and B is shown in C. Fluorescence from DAPI (D) and RFP (E) in the nucleus of a cell expressing a RFP:PYDV-N protein. The overlay of D and E is shown in F. Although both PYDV-N and -P proteins are entirely localized to the nucleus, analysis of their primary structure failed to identify karyophillic domains. G to J, Expression of AFP fusions in callus cells of N. benthamiana. G, GFP fluorescence of transgenic callus cell expressing mGFP5-ER. H, Overlay of G and I. I, RFP fluorescence following agromediated expression of RFP-SYNV-P from a pSITE vector. J, Differential interference contrast image of cell shown in G to I. K to V, Expression to study differential protein localization in pathogen-infected cells. Shown are confocal micrographs of RFP fusions of SYNV proteins expressed in SYNV-infected and mock-inoculated mGFP5-ER transgenic N. benthamiana plants. Fluorescence images for GFP, RFP, and the corresponding overlay are shown for each fusion expressed in SYNV-infected cells. Only the overlay is shown for fusions expressed in mock-inoculated leaves. Sections from top to bottom show localization of RFP:P (K–N), RFP:N (O–R), and RFP:M (S–V). Sections K to V are reprinted from Goodin et al. (2007).
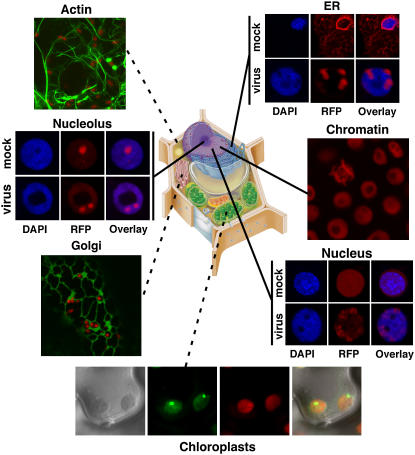
Confocal micrographs showing localization of AFPs targeted to a variety of subcellular loci. All markers are expressed from pSITE vectors in N. benthamiana. Micrographs marked with dashed lines represent transient expression while solid lines represent stable expression in transgenic plants. Plants expressing a RFP:SYNV-M fusion were not resistant to infection. Instead, RFP:SYNV-M was relocalized from the nucleoplasm to foci consistent with sites where intranuclear membranes accumulate (see Fig. 2). Clockwise, cell marker proteins used were the Rubisco small subunit (chloroplast), soybean (Glycine max) mannosidase (Golgi), Fib1 (nucleolus), RFP-HDEL (ER), Histone 2B (chromatin), and SYNV:M protein (nuclelus). Drawing of a plant cell courtesy of http://www.ualr.edu/botany/plantcelldiagram.jpg .
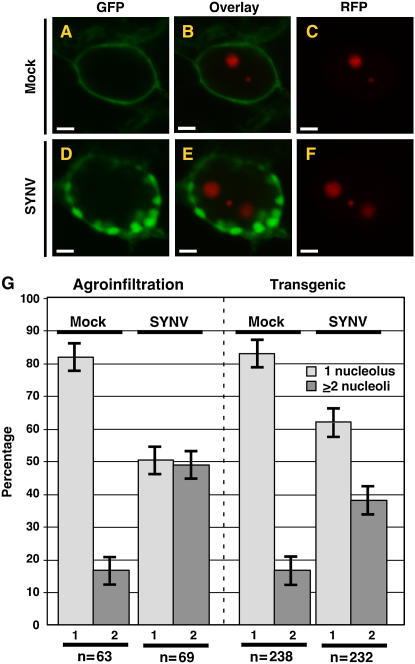
A to F, Laser scanning confocal micrographs of N. benthamiana leaf epidermal cells showing shift in the localization of expression patterns of the RFP:AtFib1 nucleolar marker from two loci in mock-inoculated cells (A–C; mock) to three in SYNV-infected cells (D–F; SYNV). Transient expression of RFP:AtFib1 was conducted in mGFP5-ER plants. G, Quantitative comparison of RFP:AtFib1 expression patterns in mock-inoculated and SYNV-infected cells. Expression patterns were divided into two categories: nuclei with one nucleolus (1, light gray), or two nucleoli (2, dark gray). Results obtained using agroinfiltration are shown on the left while those obtained with transgenic plants are on the right. The numbers of nuclei examined (n) are shown at the bottom of the graph. Sections A to F and agroinfiltration data in G are reprinted from Chakrabarty et al. (2007).
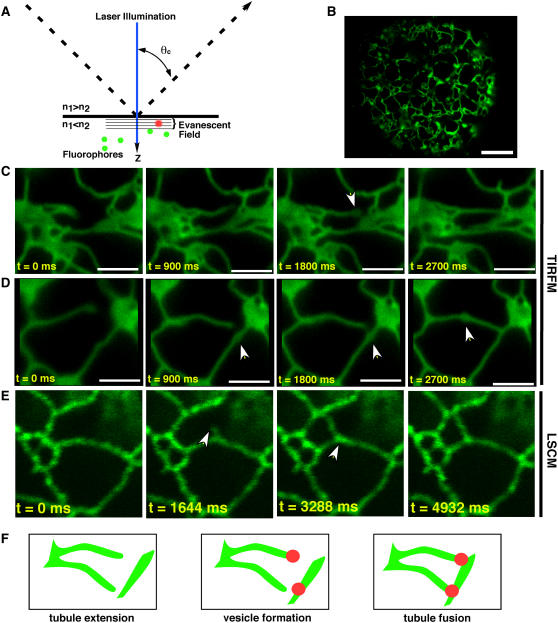
A, The underlying principle of TIRFM is that incident light rays (dashed line) that strike the interface of two media of differing refractive indices (n) at greater than the critical angle (θc) result in the rays being totally internally reflected instead of passing through the second medium. At the point of reflection an evanescent wave is produced in the medium of lower refractive index. Fluorophores (green dots) entering the evanescent wave, which is in the order of 100 nm, are excited (red dots), resulting in fluorescence detection with exceedingly high signal to noise ratios. B, TIRF micrograph of an N. benthamiana protoplast expressing m5GFP-ER. C and D, TIRF micrographs showing time series of ER tubule extension and fusion. Note that the point of fusion occurs at loci where puncta form at the termini (C) or within (D) tubules. E, Confocal micrographs showing ER tubule extension and fusion lack the speed and resolution of TIRFM. F, Model suggesting that puncta form and define the sites of ER tubule fusion.
Similar articles
-
Pumilio-based RNA in vivo imaging.Methods Mol Biol. 2015;1217:295-328. doi: 10.1007/978-1-4939-1523-1_20. Methods Mol Biol. 2015. PMID: 25287212
-
Very bright orange fluorescent plants: endoplasmic reticulum targeting of orange fluorescent proteins as visual reporters in transgenic plants.BMC Biotechnol. 2012 May 3;12:17. doi: 10.1186/1472-6750-12-17. BMC Biotechnol. 2012. PMID: 22554231 Free PMC article.
-
PSITE vectors for stable integration or transient expression of autofluorescent protein fusions in plants: probing Nicotiana benthamiana-virus interactions.Mol Plant Microbe Interact. 2007 Jul;20(7):740-50. doi: 10.1094/MPMI-20-7-0740. Mol Plant Microbe Interact. 2007. PMID: 17601162
-
In vivo RNA labeling using MS2.Methods Mol Biol. 2015;1217:329-41. doi: 10.1007/978-1-4939-1523-1_21. Methods Mol Biol. 2015. PMID: 25287213
-
GFP is the way to glow: bioimaging of the plant endomembrane system.J Microsc. 2004 May;214(Pt 2):138-58. doi: 10.1111/j.0022-2720.2004.01334.x. J Microsc. 2004. PMID: 15102062 Review.
Cited by
-
Distinctive interactions of the Arabidopsis homolog of the 30 kD subunit of the cleavage and polyadenylation specificity factor (AtCPSF30) with other polyadenylation factor subunits.BMC Cell Biol. 2009 Jul 2;10:51. doi: 10.1186/1471-2121-10-51. BMC Cell Biol. 2009. PMID: 19573236 Free PMC article.
-
Identification and Characterisation CRN Effectors in Phytophthora capsici Shows Modularity and Functional Diversity.PLoS One. 2013;8(3):e59517. doi: 10.1371/journal.pone.0059517. Epub 2013 Mar 25. PLoS One. 2013. PMID: 23536880 Free PMC article.
-
Advances in fluorescent protein-based imaging for the analysis of plant endomembranes.Plant Physiol. 2008 Aug;147(4):1469-81. doi: 10.1104/pp.108.120147. Plant Physiol. 2008. PMID: 18678739 Free PMC article. Review. No abstract available.
-
Evolutionary Analysis of Snf1-Related Protein Kinase2 (SnRK2) and Calcium Sensor (SCS) Gene Lineages, and Dimerization of Rice Homologs, Suggest Deep Biochemical Conservation across Angiosperms.Front Plant Sci. 2017 Apr 5;8:395. doi: 10.3389/fpls.2017.00395. eCollection 2017. Front Plant Sci. 2017. PMID: 28424709 Free PMC article.
-
Identification and functional assay of the interaction motifs in the partner protein OsNAR2.1 of the two-component system for high-affinity nitrate transport.New Phytol. 2014 Oct;204(1):74-80. doi: 10.1111/nph.12986. Epub 2014 Aug 7. New Phytol. 2014. PMID: 25103875 Free PMC article.
References
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301 653–657 - PubMed
-
- Bates IR, Wiseman PW, Hanrahan JW (2006) Investigating membrane protein dynamics in living cells. Biochem Cell Biol 84 825–831 - PubMed
-
- Brandizzi F, Irons SL, Johansen J, Kotzer A, Neumann U (2004) GFP is the way to glow: bioimaging of the plant endomembrane system. J Microsc 214 138–158 - PubMed
-
- Brasch MA, Hartley JL, Vidal M (2004) ORFeome cloning and systems biology: standardized mass production of the parts from the parts-list. Genome Res 14 2001–2009 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
