

The source of afterdischarge activity in neocortical tonic-clonic epilepsy
- PMID: 18057209
- PMCID: PMC6673106
- DOI: 10.1523/JNEUROSCI.3005-07.2007
The source of afterdischarge activity in neocortical tonic-clonic epilepsy
Abstract
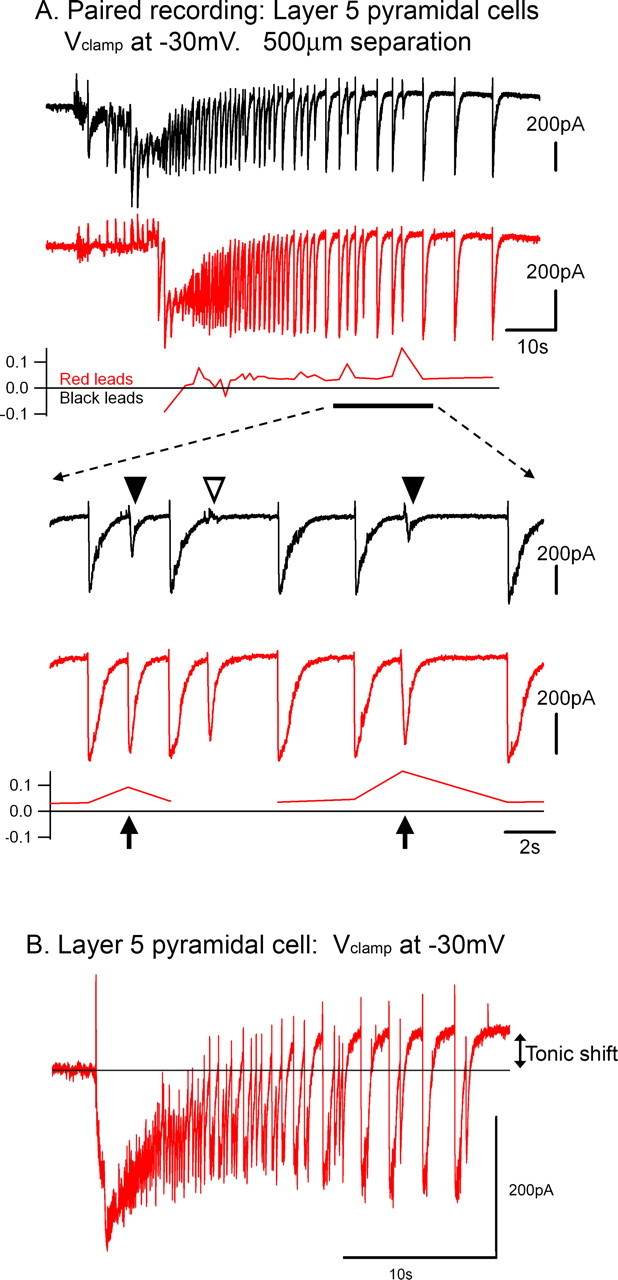
Tonic-clonic seizures represent a common pattern of epileptic discharges, yet the relationship between the various phases of the seizure remains obscure. Here we contrast propagation of the ictal wavefront with the propagation of individual discharges in the clonic phase of the event. In an in vitro model of tonic-clonic epilepsy, the afterdischarges (clonic phase) propagate with relative uniform speed and are independent of the speed of the ictal wavefront (tonic phase). For slowly propagating ictal wavefronts, the source of the afterdischarges, relative to a given recording electrode, switched as the wavefront passed by, indicating that afterdischarges are seeded from wavefront itself. In tissue that has experienced repeated ictal events, the wavefront generalizes rapidly, and the afterdischarges in this case show a different "flip-flop" pattern, with frequent switches in their direction of propagation. This same flip-flop pattern is also seen in subdural EEG recordings in patients suffering intractable focal seizures caused by cortical dysplasias. Thus, in both slowly and rapidly generalizing ictal events, there is not a single source of afterdischarge activity: rather, the source is continuously changing. Our data suggest a complex view of seizures in which the ictal event and its constituent discharges originate from distinct locations.
Conflict of interest statement
Conflict of interest statement: The authors declare no competing financial interests.
Figures
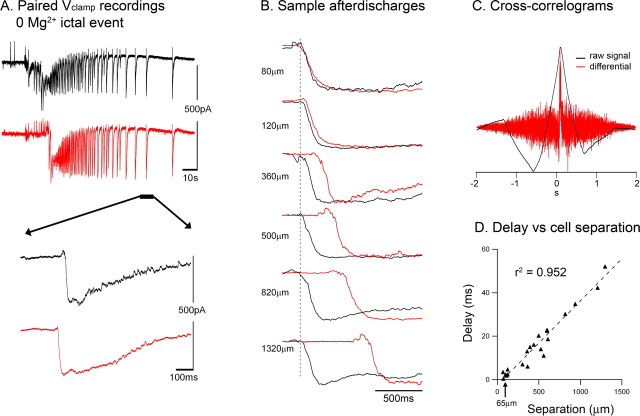

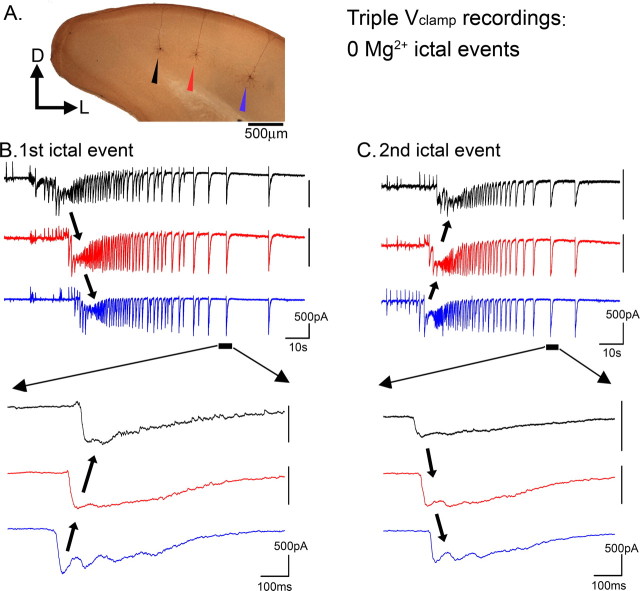
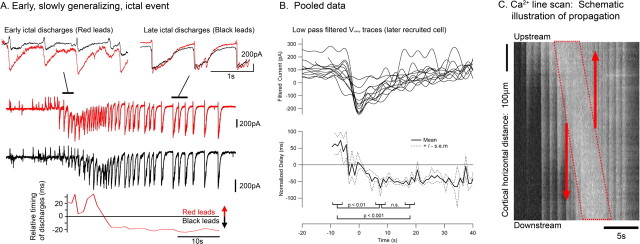
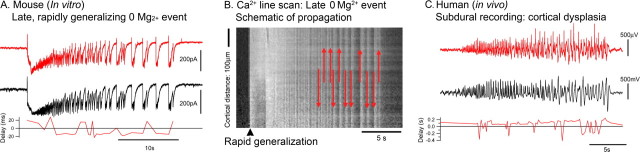
Comment in
-
Making generalizations about seizure propagation.Epilepsy Curr. 2009 Jan-Feb;9(1):24-5. doi: 10.1111/j.1535-7511.2008.01279.x. Epilepsy Curr. 2009. PMID: 19396345 Free PMC article. No abstract available.
Similar articles
-
Video-EEG evidence of lateralized clinical features in primary generalized epilepsy with tonic-clonic seizures.Epileptic Disord. 2003 Sep;5(3):149-56. Epileptic Disord. 2003. PMID: 14684350
-
Clonic convulsion caused by epileptic discharges arising from the human supplementary motor area as studied by subdural recording.Epileptic Disord. 1999 Mar;1(1):21-6. Epileptic Disord. 1999. PMID: 10937128
-
Ictal electroencephalographic findings of neonatal seizures in preterm infants.Brain Dev. 2008 Apr;30(4):261-8. doi: 10.1016/j.braindev.2007.08.011. Epub 2007 Oct 24. Brain Dev. 2008. PMID: 17920220
-
Ictal head version in generalized epilepsy.Neurology. 2004 Jul 27;63(2):370-2. doi: 10.1212/01.wnl.0000130196.79327.04. Neurology. 2004. PMID: 15277642 Review.
-
Painful generalised clonic and tonic-clonic seizures with retained consciousness.J Neurol Neurosurg Psychiatry. 1997 Dec;63(6):792-5. doi: 10.1136/jnnp.63.6.792. J Neurol Neurosurg Psychiatry. 1997. PMID: 9416819 Free PMC article. Review.
Cited by
-
Ictal wavefront propagation in slices and simulations with conductance-based refractory density model.PLoS Comput Biol. 2022 Jan 18;18(1):e1009782. doi: 10.1371/journal.pcbi.1009782. eCollection 2022 Jan. PLoS Comput Biol. 2022. PMID: 35041661 Free PMC article.
-
Field effects and ictal synchronization: insights from in homine observations.Front Hum Neurosci. 2013 Dec 5;7:828. doi: 10.3389/fnhum.2013.00828. Front Hum Neurosci. 2013. PMID: 24367311 Free PMC article. Review.
-
Excitatory effects of parvalbumin-expressing interneurons maintain hippocampal epileptiform activity via synchronous afterdischarges.J Neurosci. 2014 Nov 12;34(46):15208-22. doi: 10.1523/JNEUROSCI.1747-14.2014. J Neurosci. 2014. PMID: 25392490 Free PMC article.
-
On brain stimulation in epilepsy.Brain. 2025 Mar 6;148(3):746-752. doi: 10.1093/brain/awae385. Brain. 2025. PMID: 39745924 Free PMC article. Review.
-
Making generalizations about seizure propagation.Epilepsy Curr. 2009 Jan-Feb;9(1):24-5. doi: 10.1111/j.1535-7511.2008.01279.x. Epilepsy Curr. 2009. PMID: 19396345 Free PMC article. No abstract available.
References
-
- Alarcon G, Garcia Seoane JJ, Binnie CD, Martin Miguel MC, Juler J, Polkey CE, Elwes RD, Ortiz Blasco JM. Origin and propagation of interictal discharges in the acute electrocorticogram. Implications for pathophysiology and surgical treatment of temporal lobe epilepsy. Brain. 1997;120:2259–2282. - PubMed
-
- Albowitz B, Kuhnt U. Epileptiform activity in the guinea-pig neocortical slice spreads preferentially along supragranular layers—recordings with voltage-sensitive dyes. Eur J Neurosci. 1995;7:1273–1284. - PubMed
-
- Anderson WW, Lewis DV, Swartzwelder HS, Wilson WA. Magnesium-free medium activates seizure-like events in the rat hippocampal slice. Brain Res. 1986;398:215–219. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources