

Structural genomics of protein phosphatases
- PMID: 18058037
- PMCID: PMC4163028
- DOI: 10.1007/s10969-007-9036-1
Structural genomics of protein phosphatases
Abstract
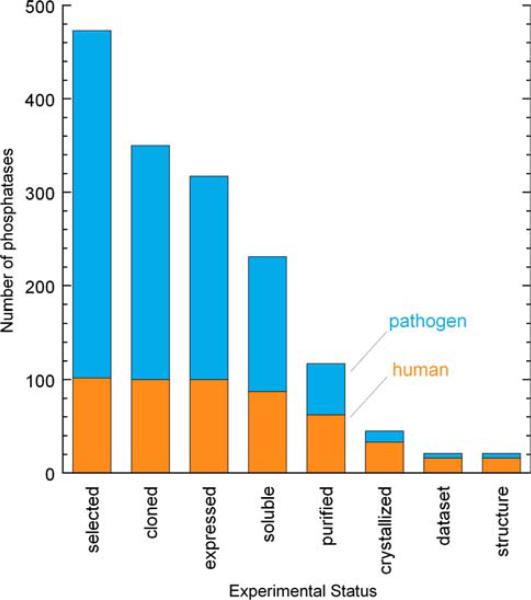
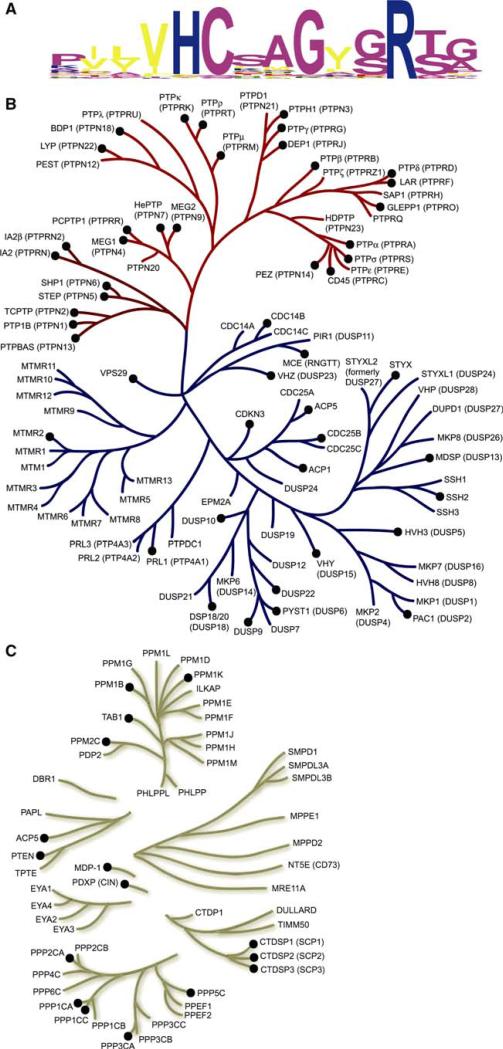
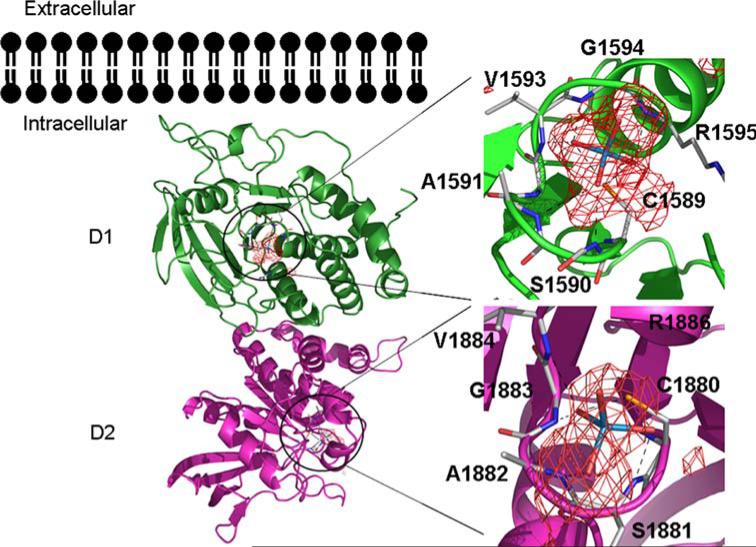
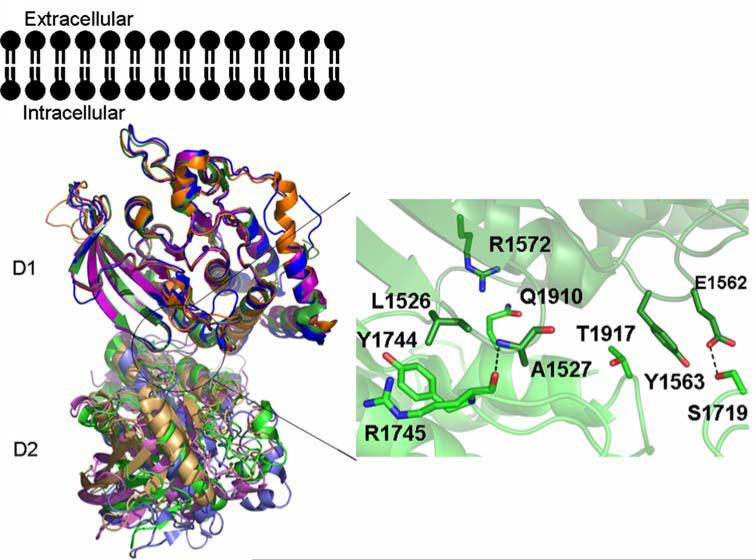
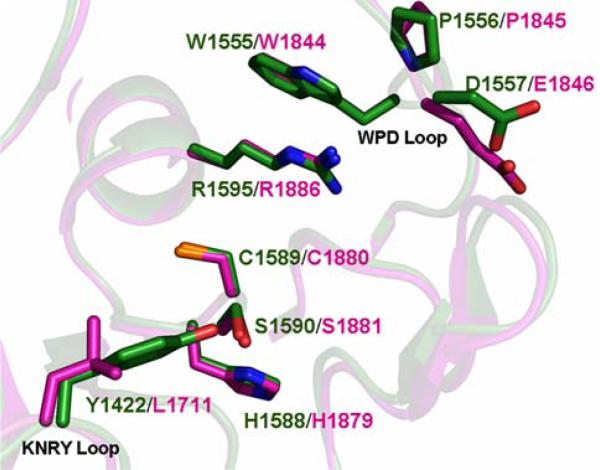
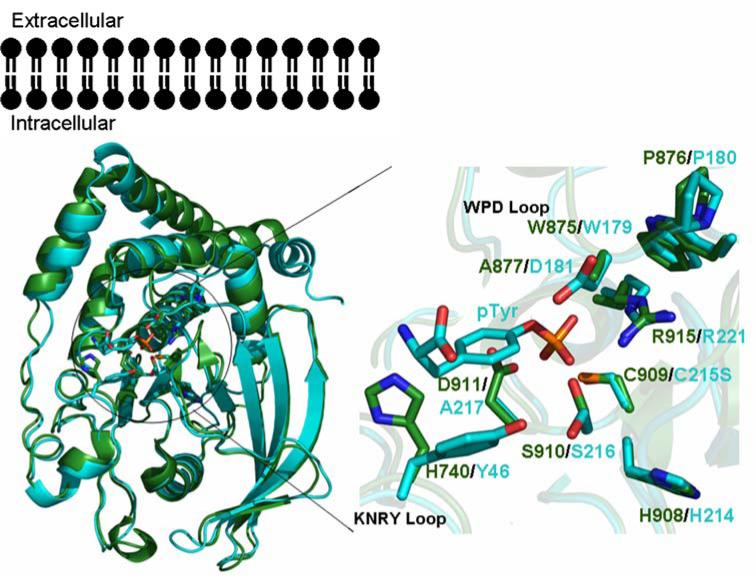
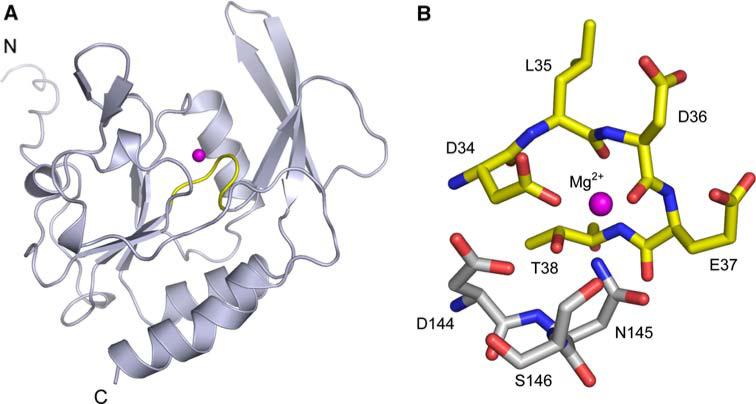
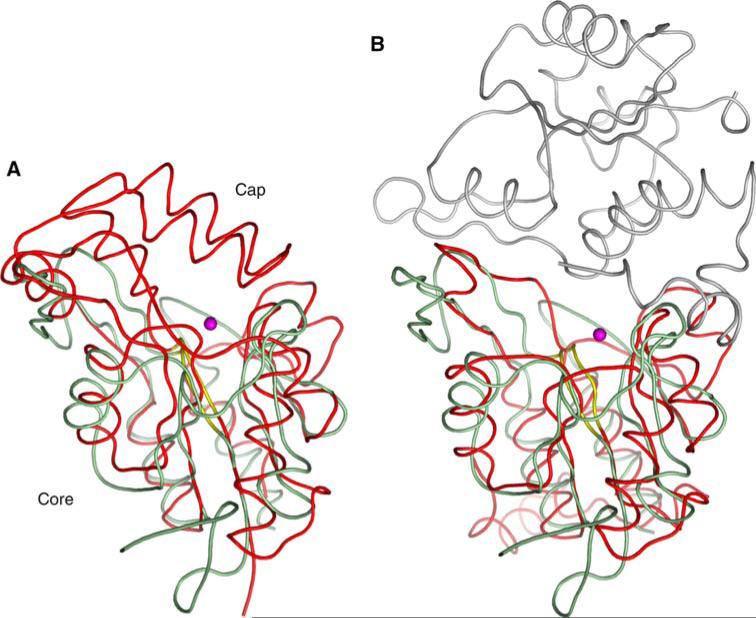
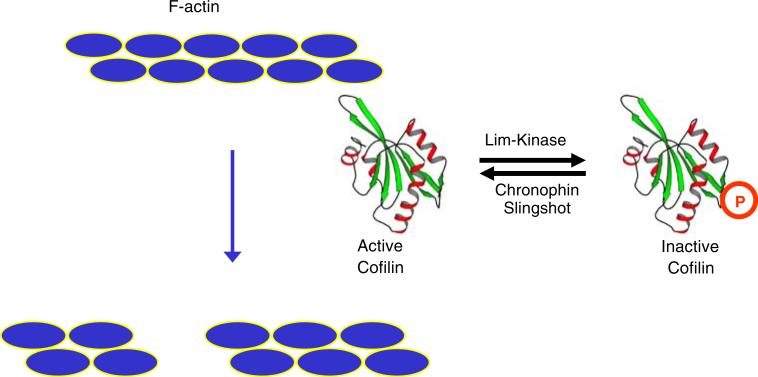
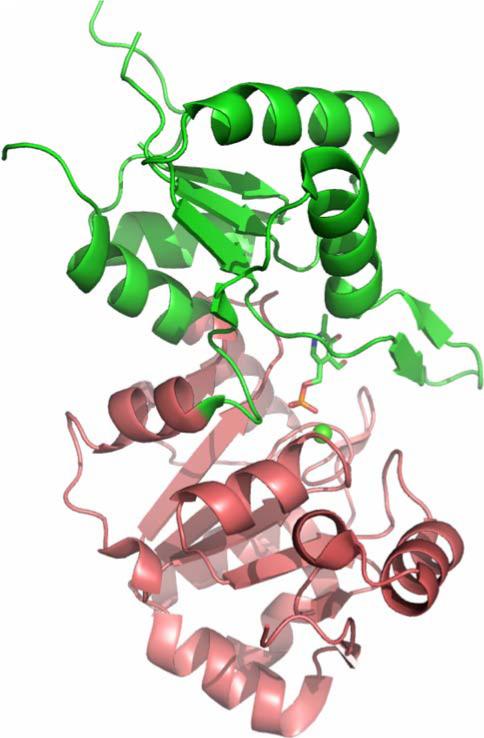
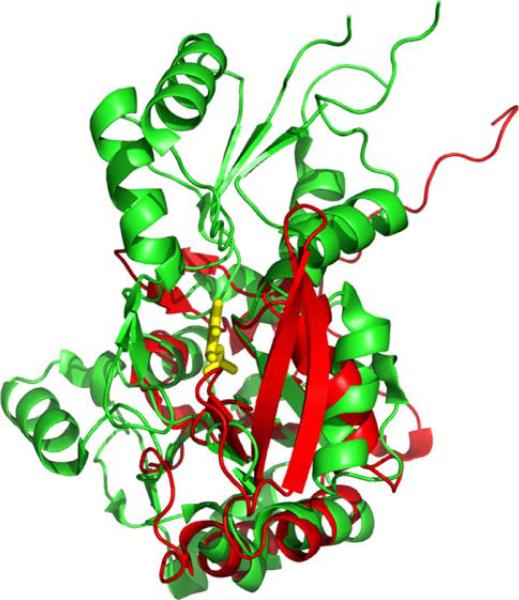
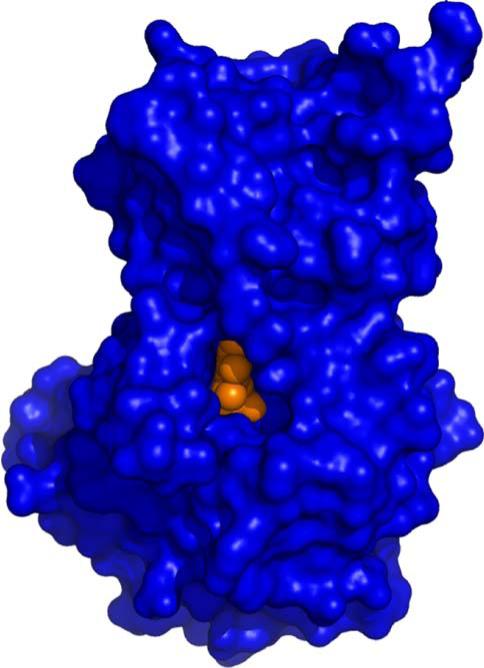
The New York SGX Research Center for Structural Genomics (NYSGXRC) of the NIGMS Protein Structure Initiative (PSI) has applied its high-throughput X-ray crystallographic structure determination platform to systematic studies of all human protein phosphatases and protein phosphatases from biomedically-relevant pathogens. To date, the NYSGXRC has determined structures of 21 distinct protein phosphatases: 14 from human, 2 from mouse, 2 from the pathogen Toxoplasma gondii, 1 from Trypanosoma brucei, the parasite responsible for African sleeping sickness, and 2 from the principal mosquito vector of malaria in Africa, Anopheles gambiae. These structures provide insights into both normal and pathophysiologic processes, including transcriptional regulation, regulation of major signaling pathways, neural development, and type 1 diabetes. In conjunction with the contributions of other international structural genomics consortia, these efforts promise to provide an unprecedented database and materials repository for structure-guided experimental and computational discovery of inhibitors for all classes of protein phosphatases.
Figures


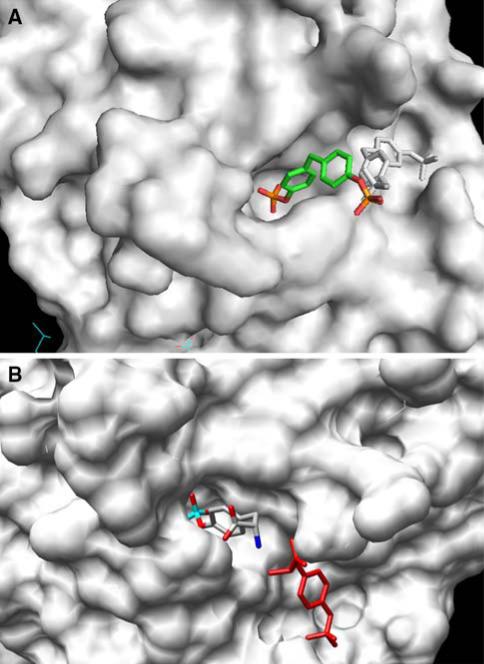
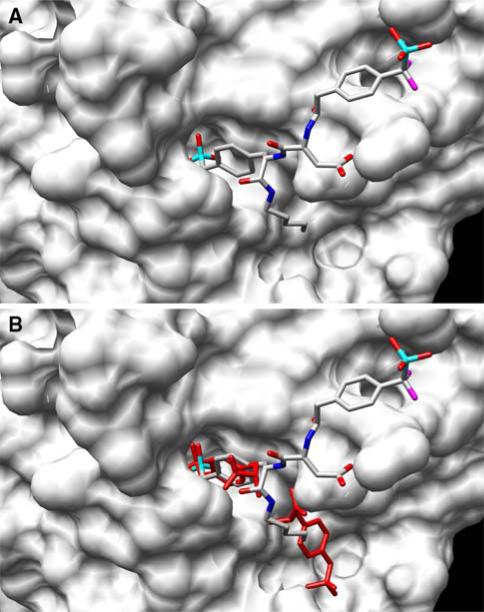
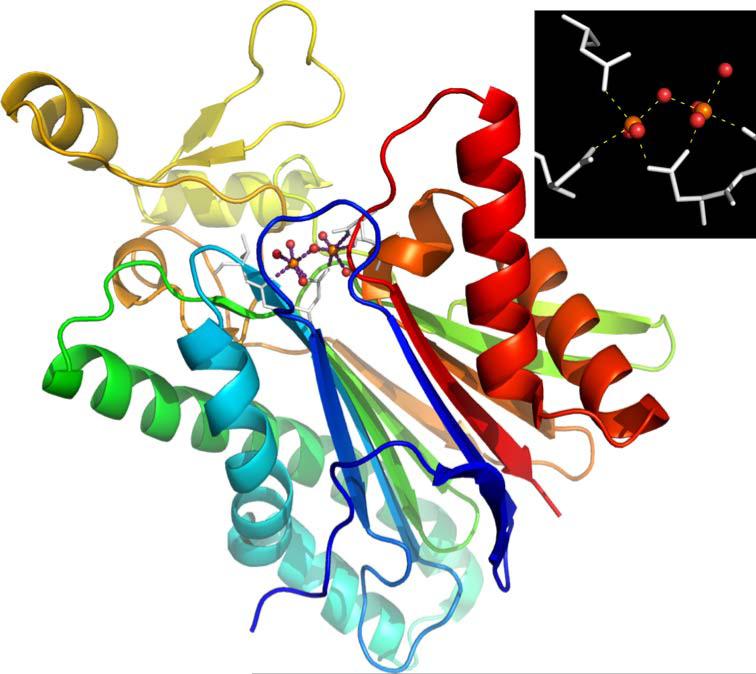
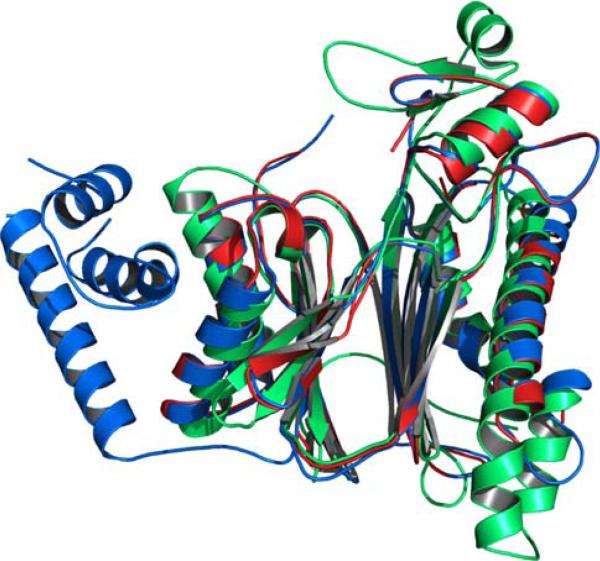
References
-
- Tonks NK. Nat Rev Mol Cell Biol. 2006;7(11):833–846. - PubMed
-
- McCluskey A, Sim AT, Sakoff JA. J Med Chem. 2002;45(6):1151–1175. - PubMed
-
- Peisach E, Selengut JD, Dunaway-Mariano D, Allen KN. Biochemistry. 2004;43(40):12770–12779. - PubMed
-
- Wiggan O, Bernstein BW, Bamburg JR. Nat Cell Biol. 2005;7(1):8–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
