

Stabilization of overlapping microtubules by fission yeast CLASP
- PMID: 18061564
- PMCID: PMC2215317
- DOI: 10.1016/j.devcel.2007.10.015
Stabilization of overlapping microtubules by fission yeast CLASP
Abstract
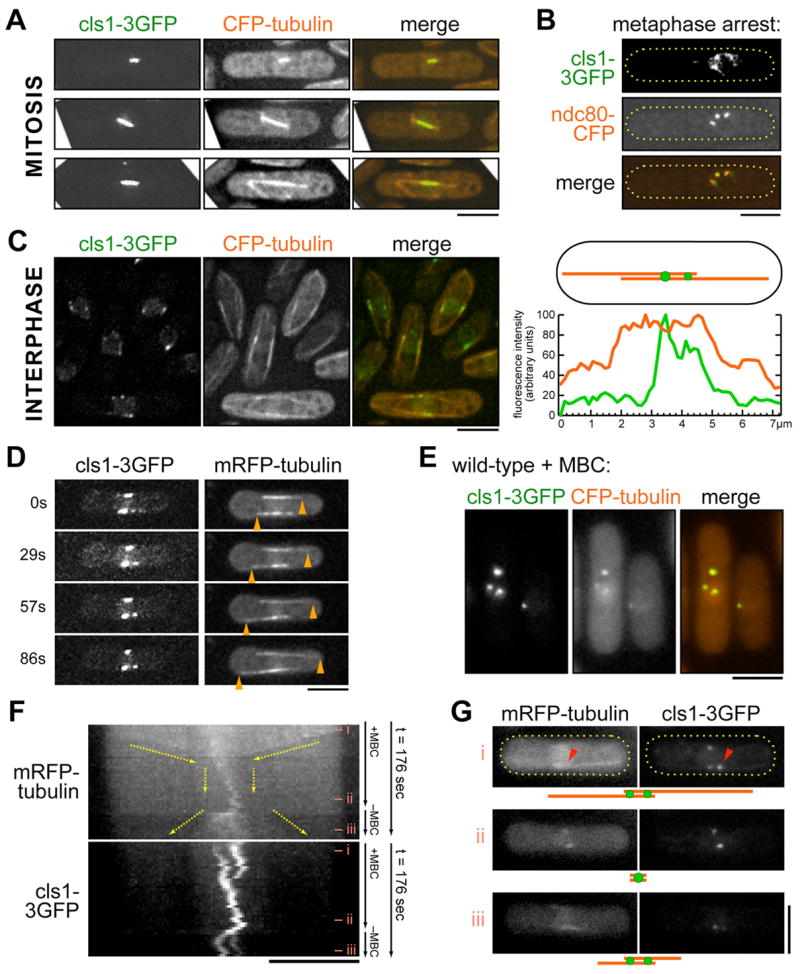
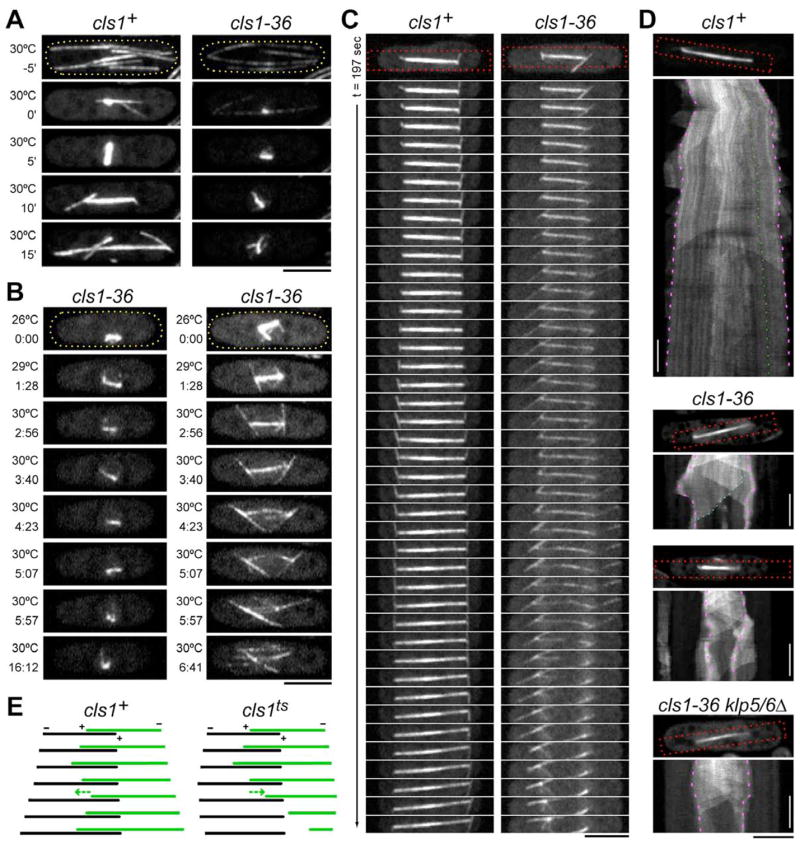
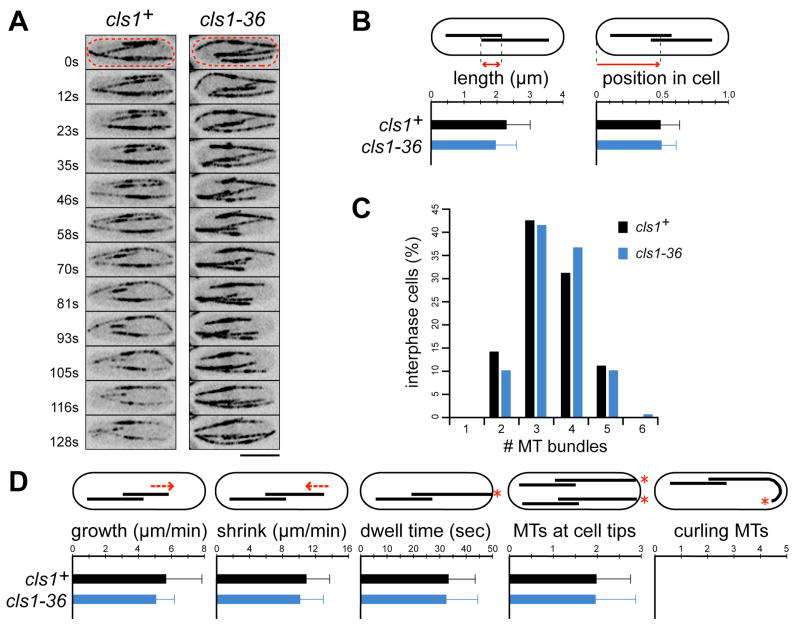
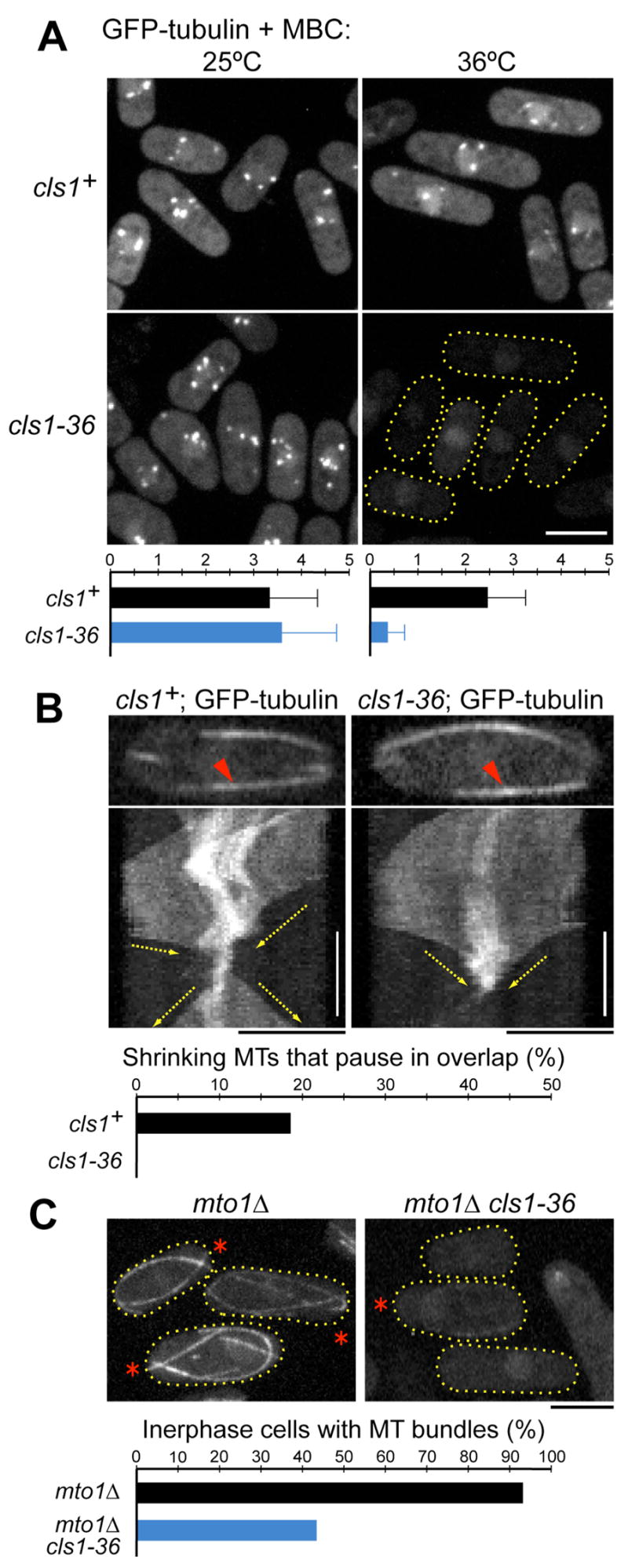
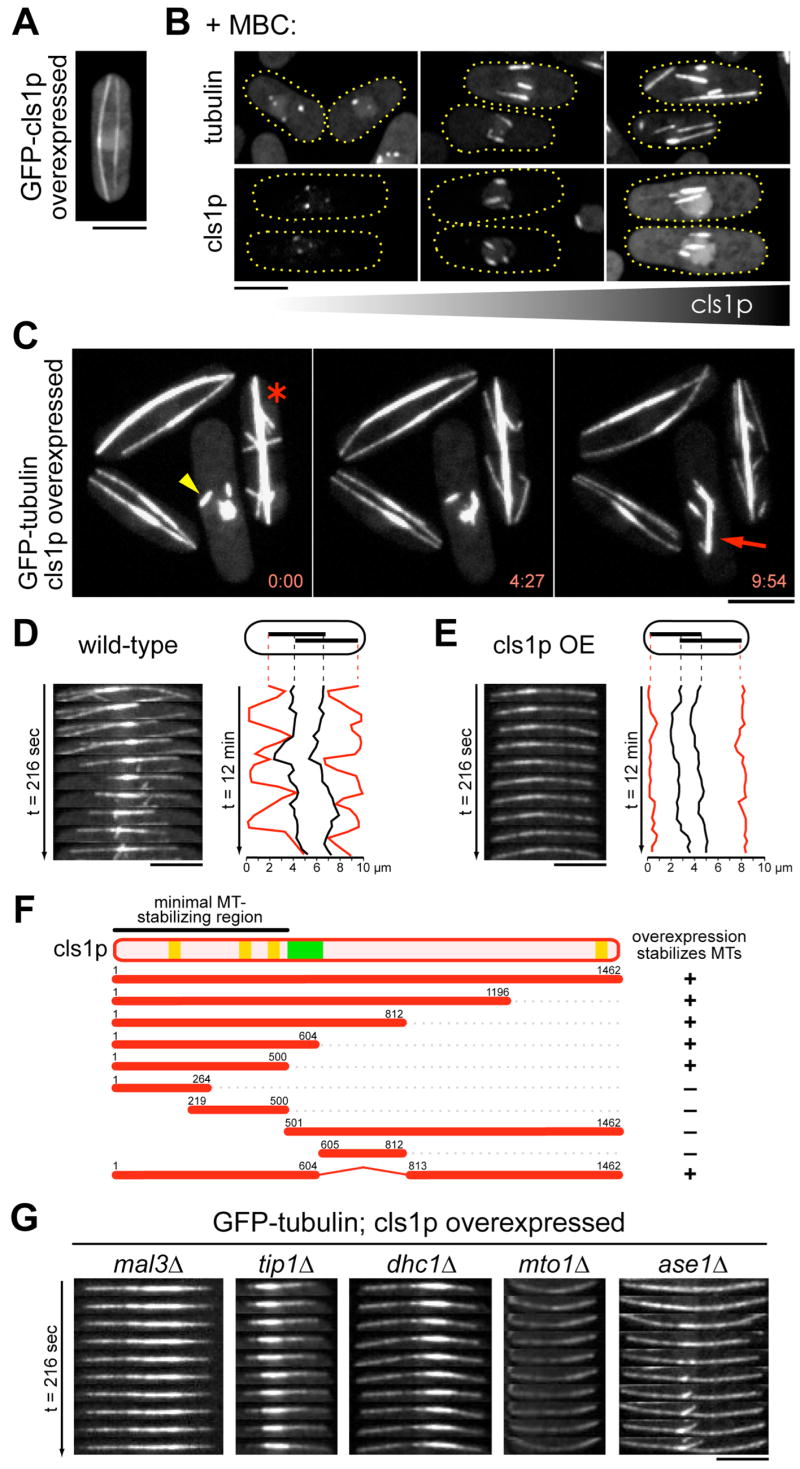
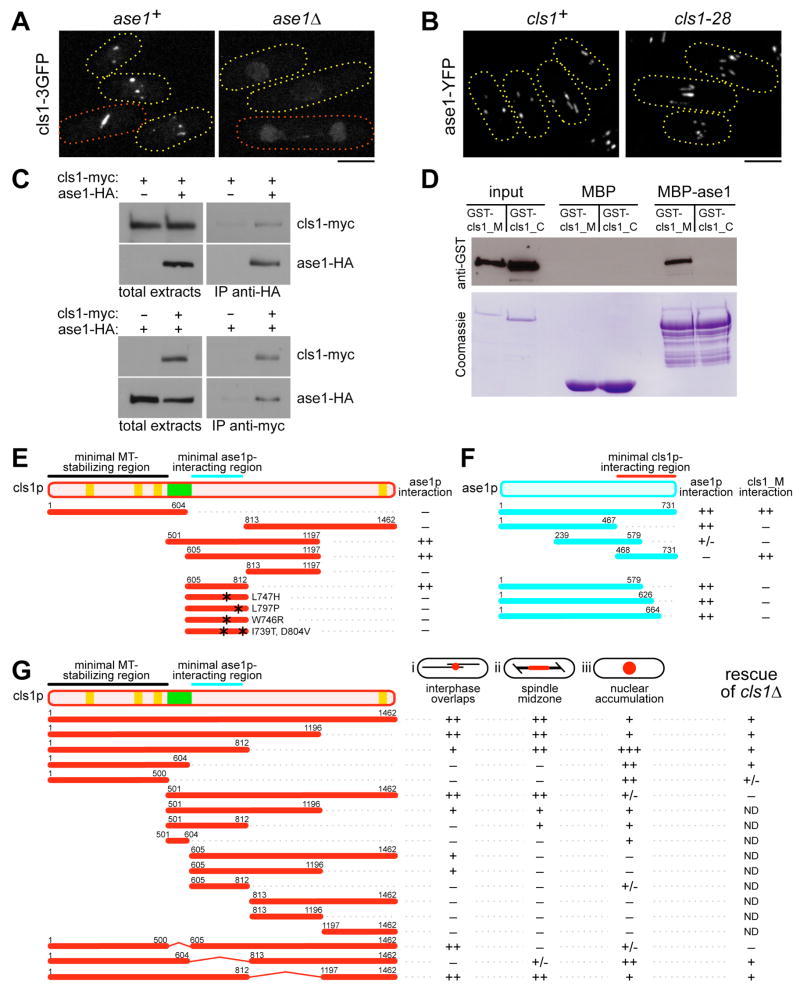
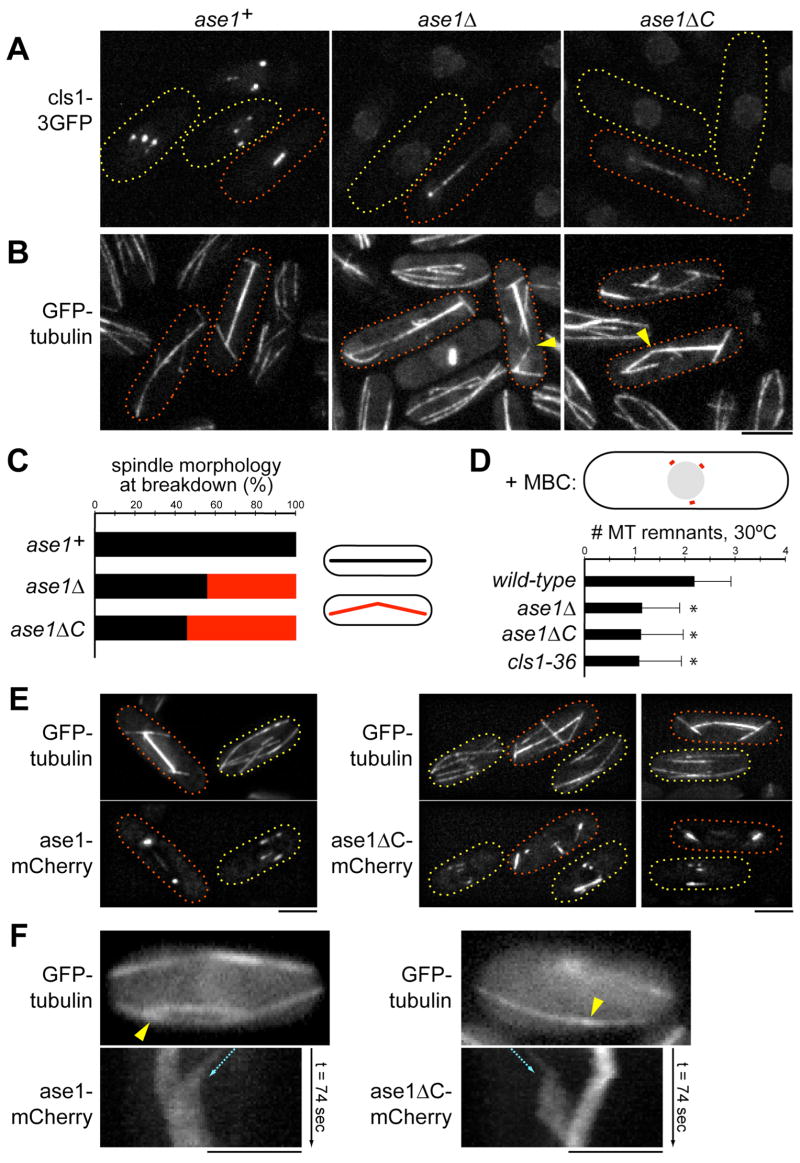
Many microtubule (MT) structures contain dynamic MTs that are bundled and stabilized in overlapping arrays. CLASPs are conserved MT-binding proteins implicated in the regulation of MT plus ends. Here, we show that the Schizosaccharomyces pombe CLASP, cls1p/peg1p, mediates the stabilization of overlapping MTs within the mitotic spindle and interphase bundles. cls1p localizes to these regions but not to interphase MT plus ends. Inactivation of cls1p leads to the rapid depolymerization of spindle midzone MTs. cls1p also stabilizes a subset of MTs within interphase bundles. cls1p prevents disassembly of the entire microtubule, while still allowing for plus-end growth. It has no measurable effects on MT nucleation, polymerization, catastrophe, or bundling. A direct interaction with ase1p (PRC1/MAP65) targets cls1p to regions of antiparallel MT overlap. These findings show how a MT-stabilizing factor attached to specific sites on MTs can help to generate MT structures that have both dynamic and stable components.
Figures
References
-
- Akhmanova A, Hoogenraad CC. Microtubule plus-end-tracking proteins: mechanisms and functions. Curr Opin Cell Biol. 2005;17:47–54. - PubMed
-
- Akhmanova A, Hoogenraad CC, Drabek K, Stepanova T, Dortland B, Verkerk T, Vermeulen W, Burgering BM, De Zeeuw CI, Grosveld F, Galjart N. Clasps are CLIP-115 and -170 associating proteins involved in the regional regulation of microtubule dynamics in motile fibroblasts. Cell. 2001;104:923–935. - PubMed
-
- Al-Bassam J, Larsen NA, Hyman AA, Harrison SC. Crystal structure of a TOG domain: conserved features of XMAP215/Dis1-family TOG domains and implications for tubulin binding. Structure. 2007;15:355–362. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
