

Biochemical activities of Arabidopsis RNA-dependent RNA polymerase 6
- PMID: 18063577
- PMCID: PMC2629599
- DOI: 10.1074/jbc.M708983200
Biochemical activities of Arabidopsis RNA-dependent RNA polymerase 6
Abstract
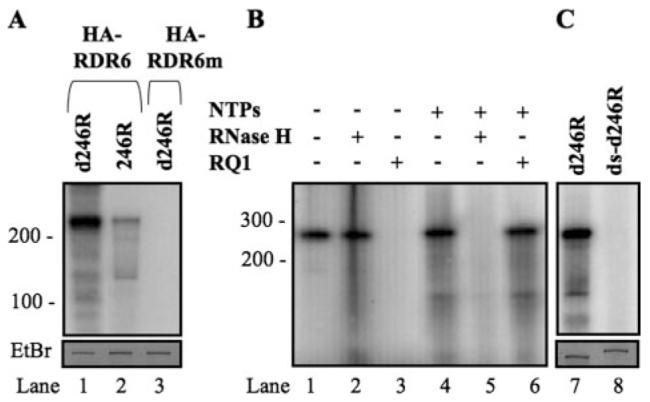
In Arabidopsis, genetic evidence demonstrates that RNA-dependent RNA polymerase 6 (RDR6) plays a fundamental role in at least four RNA silencing pathways whose functions range from defense against transgenes or viruses to endogene regulation in development and in stress responses. Despite its critical role in RNA silencing, the biochemical activities of RDR6 have yet to be characterized. In this study, we transiently expressed Arabidopsis RDR6 in Nicotiana benthamiana and investigated the biochemical activities of immunopurified RDR6 in vitro. We showed that RDR6 possesses terminal nucleotidyltransferase activity as well as primer-independent RNA polymerase activity on single-stranded RNAs. We found that RDR6 cannot distinguish RNAs with or without a cap or poly(A) tail. We also demonstrated that RDR6 has strong polymerase activity on single-stranded DNA. All these activities require the conserved catalytic Asp(867) residue. Our findings have important implications on the processes involving RDR6 in vivo and provide new biochemical insights into the mechanisms of RNA silencing in Arabidopsis.
Figures
References
-
- Peters L, Meister G. Mol. Cell. 2007;26:611–623. - PubMed
-
- Cogoni C, Macino G. Nature. 1999;399:166–169. - PubMed
-
- Dalmay T, Hamilton A, Rudd S, Angell S, Baulcombe DC. Cell. 2000;101:543–553. - PubMed
-
- Mourrain P, Beclin C, Elmayan T, Feuerbach F, Godon C, Morel JB, Jouette D, Lacombe AM, Nikic S, Picault N, Remoue K, Sanial M, Vo TA, Vaucheret H. Cell. 2000;101:533–542. - PubMed
-
- Smardon A, Spoerke JM, Stacey SC, Klein ME, Mackin N, Maine EM. Curr. Biol. 2000;10:169–178. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
