

Modulation of ileal bile acid transporter (ASBT) activity by depletion of plasma membrane cholesterol: association with lipid rafts
- PMID: 18063707
- PMCID: PMC4880014
- DOI: 10.1152/ajpgi.00237.2007
Modulation of ileal bile acid transporter (ASBT) activity by depletion of plasma membrane cholesterol: association with lipid rafts
Abstract
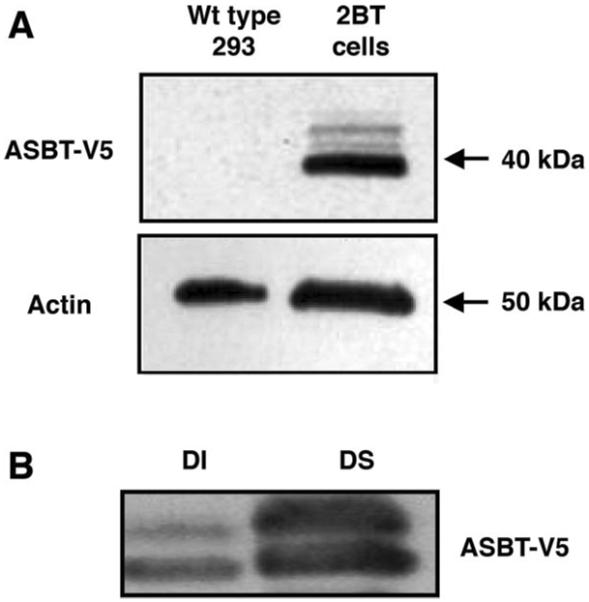
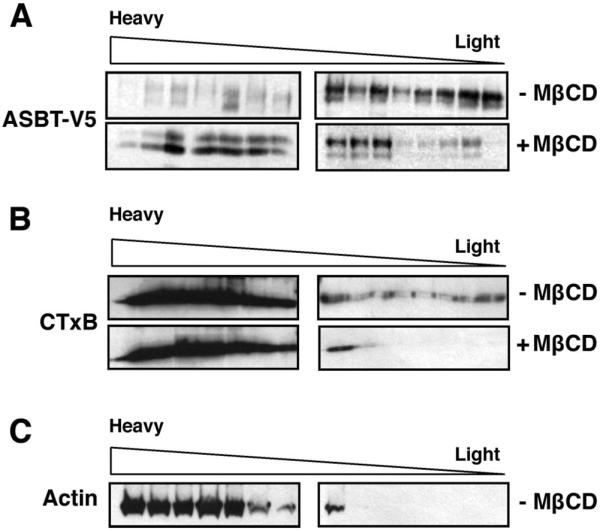
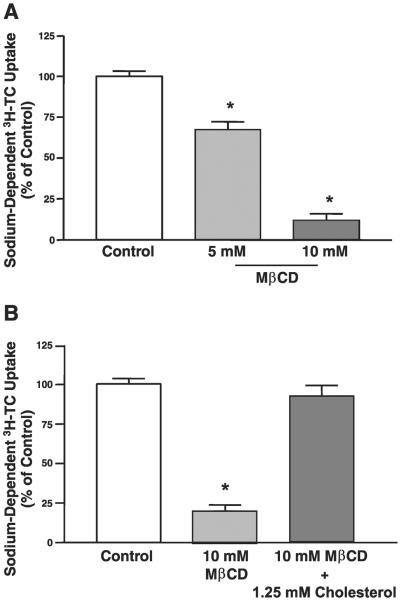
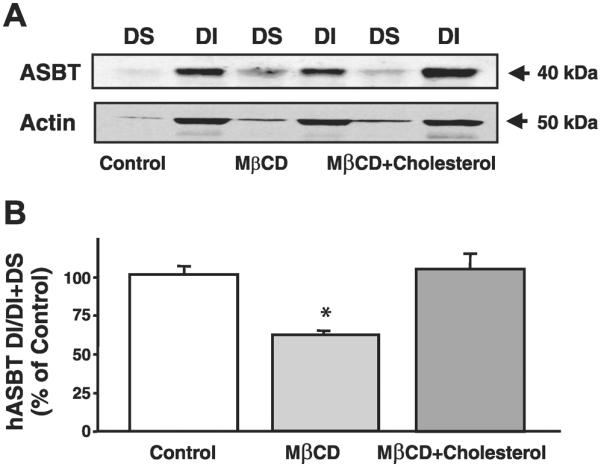
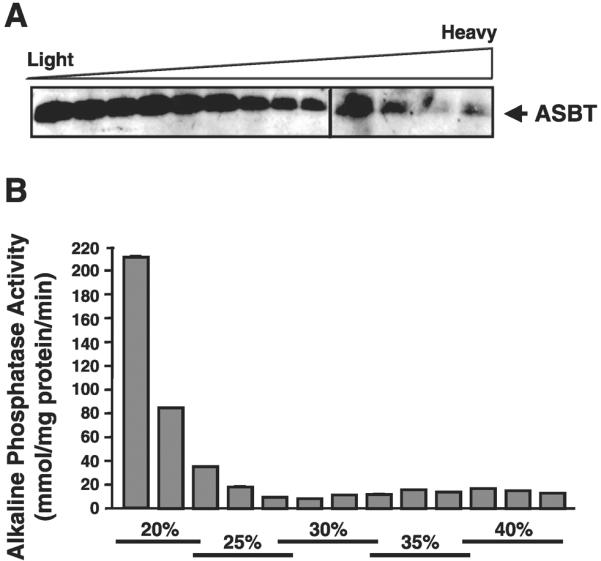
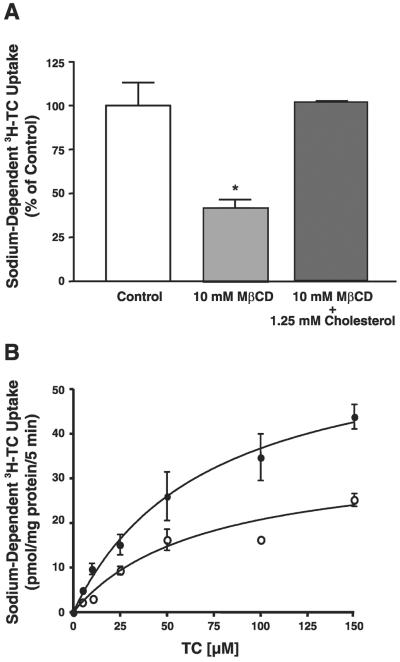
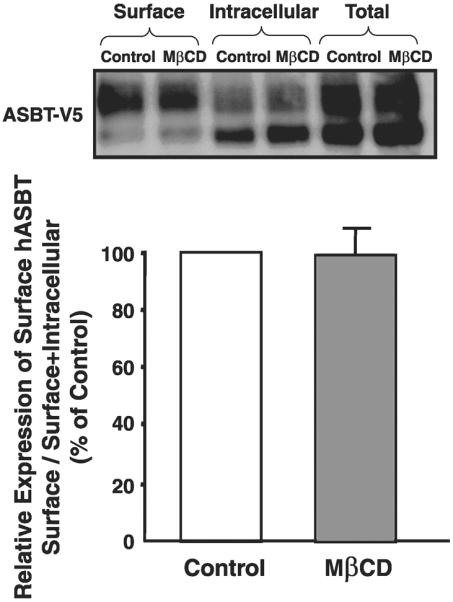
Apical sodium-dependent bile acid transporter (ASBT) represents a highly efficient conservation mechanism of bile acids via mediation of their active transport across the luminal membrane of terminal ileum. To gain insight into the cellular regulation of ASBT, we investigated the association of ASBT with cholesterol and sphingolipid-enriched specialized plasma membrane microdomains known as lipid rafts and examined the role of membrane cholesterol in maintaining ASBT function. Human embryonic kidney (HEK)-293 cells stably transfected with human ASBT, human ileal brush-border membrane vesicles, and human intestinal epithelial Caco-2 cells were utilized for these studies. Floatation experiments on Optiprep density gradients demonstrated the association of ASBT protein with lipid rafts. Disruption of lipid rafts by depletion of membrane cholesterol with methyl-beta-cyclodextrin (MbetaCD) significantly reduced the association of ASBT with lipid rafts, which was paralleled by a decrease in ASBT activity in Caco-2 and HEK-293 cells treated with MbetaCD. The inhibition in ASBT activity by MbetaCD was blocked in the cells treated with MbetaCD-cholesterol complexes. Kinetic analysis revealed that MbetaCD treatment decreased the V(max) of the transporter, which was not associated with alteration in the plasma membrane expression of ASBT. Our study illustrates that cholesterol content of lipid rafts is essential for the optimal activity of ASBT and support the association of ASBT with lipid rafts. These findings suggest a novel mechanism by which ASBT activity may be rapidly modulated by alterations in cholesterol content of plasma membrane and thus have important implications in processes related to maintenance of bile acid and cholesterol homeostasis.
Figures
References
-
- Akhter S, Cavet ME, Tse CM, Donowitz M. C-terminal domains of Na+/H+ exchanger isoform 3 are involved in the basal and serum-stimulated membrane trafficking of the exchanger. Biochemistry. 2000;39:1990–2000. - PubMed
-
- Alpini G, Glaser S, Baiocchi L, Francis H, Xia X, Lesage G. Secretin activation of the apical Na+-dependent bile acid transporter is associated with cholehepatic shunting in rats. Hepatology. 2005;41:1037–1045. - PubMed
-
- Alpini G, Glaser SS, Rodgers R, Phinizy JL, Robertson WE, Lasater J, Caligiuri A, Tretjak Z, LeSage GD. Functional expression of the apical Na+-dependent bile acid transporter in large but not small rat cholangiocytes. Gastroenterology. 1997;113:1734–1740. - PubMed
-
- Alrefai WA, Saksena S, Tyagi S, Gill RK, Ramaswamy K, Dudeja PK. Taurodeoxycholate modulates apical Cl−/OH− exchange activity in Caco2 cells. Dig Dis Sci. 2007;52:1270–1278. - PubMed
-
- Alrefai WA, Sarwar Z, Tyagi S, Saksena S, Dudeja PK, Gill RK. Cholesterol modulates human intestinal sodium-dependent bile acid transporter. Am J Physiol Gastrointest Liver Physiol. 2005;288:G978–G985. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P01 DK067887/DK/NIDDK NIH HHS/United States
- DK-68271/DK/NIDDK NIH HHS/United States
- DK-71596/DK/NIDDK NIH HHS/United States
- P01 DK-067887/DK/NIDDK NIH HHS/United States
- R01 DK054016/DK/NIDDK NIH HHS/United States
- R01 DK061931/DK/NIDDK NIH HHS/United States
- R01 DK068271/DK/NIDDK NIH HHS/United States
- R56 DK071596/DK/NIDDK NIH HHS/United States
- R01 DK068324/DK/NIDDK NIH HHS/United States
- R01 DK071596/DK/NIDDK NIH HHS/United States
- R56 DK094954/DK/NIDDK NIH HHS/United States
- DK-68324/DK/NIDDK NIH HHS/United States
- DK-54016/DK/NIDDK NIH HHS/United States
- DK-61931/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous